Th17 cells in organ transplantation

127
Th 17 Cells: Role in Inflammation and Autoimmune Disease,
edited by Valérie Quesniaux, Bernhard Ryffel and Franco Di Padova
? 2009 Birkh?user Verlag Basel/Switzerland Th17 cells in organ transplantation
Ye Chen 1 and Kathryn J. Wood 2
1
Sir William Dunn School of Pathology, University of Oxford, Oxford, UK; 2Transplantation Research Immunology Group, Nuffield Department of Surgery, University of Oxford, Oxford, UK
Abstract
A newly discovered subset of T helper cells, Th17 cells, has been implicated in a number of models of autoimmunity. However, their role in transplantation remains undefined. This is despite studies showing a link between IL-17, a cytokine secreted by Th17 cells, and acute allograft rejection. This chapter summarises current evidence for a role of Th17 cells in transplant rejection, tolerance and ischaemia reperfusion injury.
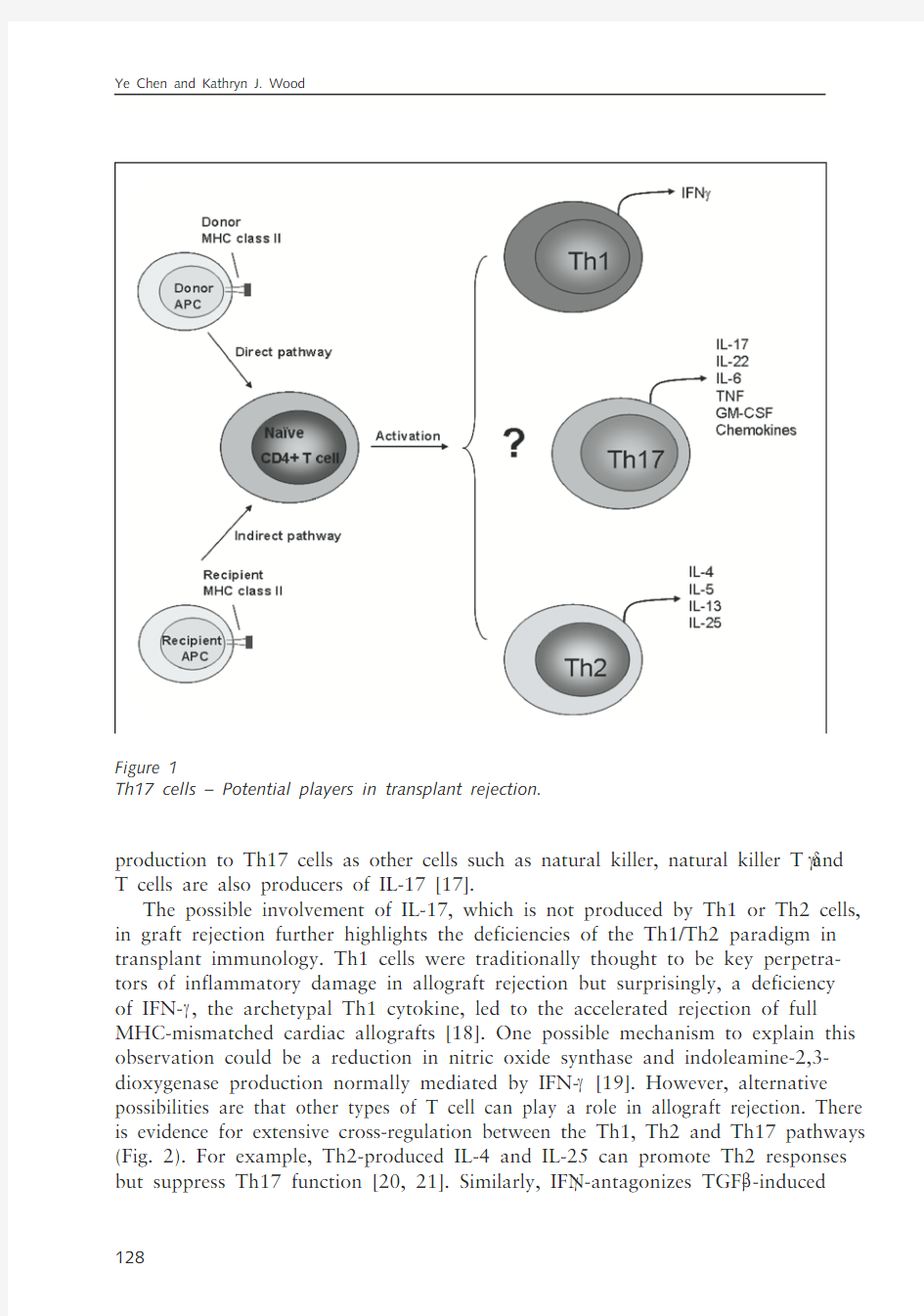
Th17 cells are a recently discovered ‘arm’ of T helper cells that can differentiate from na?ve T helper cells following their activation. Although Th17 cells have been cast as major instigators of inflammatory damage in a number of models of autoim-munity, including collagen-induced arthritis (CIA) [1–3], experimental autoimmune encephalomyelitis (EAE) [4–7] and dermal inflammation [8], their role in transplan-tation immunity is ambiguous (Fig. 1). A recent study suggests that donor-derived Th17 and Th1 cells can mediate chronic graft-versus -host-disease via enhanced autoreactivity [9]. In solid-organ rejection models, some studies have demonstrated a link between the Th17-secreted cytokine IL-17 and acute allograft rejection.
IL-17 is a proinflammatory cytokine that has multiple targets and triggers the release of further mediators downstream. Predominantly, it functions to stimulate granulopoiesis and the migration of neutrophils to the site of inflammation [10]. In one study designed to document the kinetics of IL-17 mRNA expression during renal allograft rejection in a rat model, IL-17 mRNA was detectable by postopera-tive day 2, peaked at day 5 before declining, becoming undetectable by day 9 [11]. Neutralization of IL-17 by administration of an IL-17R:IgG-Fc fusion protein was found able to prevent the acute rejection of aortic allografts [12] and to prolong the survival of cardiac allografts [13, 14]. In humans, increased IL-17 protein levels have been detected in rejecting renal allografts and in bronchoalveolar lavage dur-ing acute lung allograft rejection [15, 16]. Further studies are needed to link IL-17
128
production to Th17 cells as other cells such as natural killer, natural killer T and GD T cells are also producers of IL-17 [17].
The possible involvement of IL-17, which is not produced by Th1 or Th2 cells, in graft rejection further highlights the deficiencies of the Th1/Th2 paradigm in transplant immunology. Th1 cells were traditionally thought to be key perpetra-tors of inflammatory damage in allograft rejection but surprisingly, a deficiency of IFN-G , the archetypal Th1 cytokine, led to the accelerated rejection of full MHC-mismatched cardiac allografts [18]. One possible mechanism to explain this observation could be a reduction in nitric oxide synthase and indoleamine-2,3-dioxygenase production normally mediated by IFN-G [19]. However, alternative possibilities are that other types of T cell can play a role in allograft rejection. There is evidence for extensive cross-regulation between the Th1, Th2 and Th17 pathways (Fig. 2). For example, Th2-produced IL-4 and IL-25 can promote Th2 responses but suppress Th17 function [20, 21]. Similarly, IFN-G antagonizes TGF-B
-induced Figure 1
Th17 cells – Potential players in transplant rejection.
Figure 2
Cross-regulation between Th1, Th2 and Th17 pathways.
Th17 development and inhibits IL-17 and IL-23 production [20]. Allograft pathol-ogy in IFN-G-deficient animals showed reduced mononuclear infiltration and more intense neutrophil infiltration [18]. This may arise as a consequence of the lifting of the inhibitory signal on Th17 differentiation by the absence of IFN-G, leading to increased IL-17 production and neutrophil recruitment.
Th17 cells may also share a complex link with regulatory T cells (Tregs), with consequent ramifications for the potential use of Tregs to induce transplant tolerance [22]. When exposed to TGF-B, na?ve mouse CD4+ T cells (nTh) can differentiate into Tregs [23]. However, nTh can develop into Th17 cells when exposed to TGF-B and IL-6 [24]. Therefore, in this setting, IL-6 appears to be a ‘switch’ factor that determines whether nTh acquires a regulatory or a proinflammatory phenotype.
In vitro, IL-6 and TGF-B-producing Tregs can generate IL-17-producing T cells from nTh [25, 26]. Furthermore, a recent study showed that Tregs, in the pres-ence of IL-6 or activated dendritic cells, can themselves differentiate into IL-17-producing Th17 cells [26]. In a T cell-mediated autoimmunity model, adoptively transferred Tregs increased IL-17 production by effector T cells and failed to prevent disease established by these cells [27]. Kuchroo and colleagues [7] also showed that, in EAE, Tregs were unable to abrogate IL-17 production by T cells that produced high amounts of IL-6 and TNF-A. In an inflammatory milieu, IL-6 production may tip the balance against Treg regulation towards Th17 induction. In transplantation,
129
such an inflammatory milieu may be established by ischaemia reperfusion injury (IRI) of the transplanted graft.
IRI in the transplant setting is defined as the period from when the graft is retrieved from the donor (ischaemia) to sometime after its revascularisation in the recipient (reperfusion) [28]. The ischaemic phase may include a ‘warm ischaemic interval’ before or during removal from the donor, and a ‘cold ischemic interval’ associated with preservation and storage of the graft. IRI contributes to acute graft failure, delayed graft function and increased alloimmunity and graft rejection [29]).
IL-6, an essential differentiation factor for Th17 cells, is implicated in IRI. IL-6 is expressed within 1 h of reperfusion following renal ischaemia [30]. Urinary IL-6 levels also appear to correlate with the severity of renal injury [31]. Both mice that have been treated with anti-IL-6 antibody or IL-6-deficient mice show attenu-ated renal dysfunction arising from IRI and have reduced neutrophil infiltration and expression of adhesion molecules [32]. In renal IRI, macrophages may be a source of IL-6. The transfer of IL-6-sufficient macrophages into IL-6-deficient mice restores susceptibility to IRI [33]. Increased IL-6 expression in IRI may lead to the induction of Th17 cells. Recently, Yoshida and colleagues [34] have shown that lung ischaemia reperfusion injury could induce type V collagen-specific T cells that secreted IL-17.
The role of Th17 cells is still unclear in transplantation immunology. Along with Th1 cells, Th17 cells may mediate allograft rejection and their role may be more important when Th1 responses are suppressed. Their relationship with Tregs in the transplantation setting warrants further investigation as it may mean anti-IL-6 agents are needed to block Th17 differentiation to enable effective Treg function in establishing transplant tolerance. Finally, IL-6 induced by IRI may promote IL-17 production and neutrophil infiltration, thus contributing to acute graft failure. References
1 Nakae S, Nambu A, Sudo K, Iwakura Y. Suppression of immune induction of collagen-
induced arthritis in IL-17-deficient mice. J Immunol 2003; 171: 6173–6177
2 Hirota K, Hashimoto M, Yoshitomi H et al. T cell self-reactivity forms a cytokine milieu
for spontaneous development of IL-17+ Th cells that cause autoimmune arthritis. J Exp Med 2007; 204: 41–47
3 Sato K, Suematsu A, Okamoto K et al. Th17 functions as an osteoclastogenic helper
T cell subset that links T cell activation and bone destruction. J Exp Med 2006; 203: 2673–2682
4 Cua DJ, Sherlock J, Chen Y et al. Interleukin-23 rather than interleukin-12 is the critical
cytokine for autoimmune inflammation of the brain. Nature 2003; 421: 744–748
130
5 Park H, Li Z, Yang XO et al. A distinct lineage of CD4 T cells regulates tissue inflam-
mation by producing interleukin 17. Nat Immunol 2005; 6: 1133–1141
6 Batten M, Li J, Yi S et al. Interleukin 2
7 limits autoimmune encephalomyelitis by sup-
pressing the development of interleukin 17-producing T cells. Nat Immunol 2006; 7: 929–936
7 Korn T, Reddy J, Gao W et al. Myelin-specific regulatory T cells accumulate in the CNS
but fail to control autoimmune inflammation. Nat Med 2007; 13: 423–431
8 Zheng Y, Danilenko DM, Valdez P et al. Interleukin-22, a T(H)17 cytokine, mediates
IL-23–induced dermal inflammation and acanthosis. Nature 2007; 445: 648–651
9 Chen X, Vodanovic-Jankovic S, Johnson B, Keller M, Komorowski R, Drobyski WR.
Absence of regulatory T-cell control of TH1 and TH17 cells is responsible for the autoimmune-mediated pathology in chronic graft-versus-host disease. Blood 2007; 110: 3804–3813
10 Witowski J, Ksiazek K, Jorres A. Interleukin-17: A mediator of inflammatory responses.
Cell Mol Life Sci 2004; 61: 567–579
11 Loong CC, Hsieh HG, Lui WY, Chen A, Lin CY. Evidence for the early involvement of
interleukin 17 in human and experimental renal allograft rejection. J Pathol 2002; 197: 322–332
12 Tang JL, Subbotin VM, Antonysamy MA, Troutt AB, Rao AS, Thomson AW. Interleu-
kin-17 antagonism inhibits acute but not chronic vascular rejection. Transplantation 2001; 72: 348–350
13 Antonysamy MA, Fanslow WC, Fu F et al. Evidence for a role of IL-17 in organ allograft
rejection: IL-17 promotes the functional differentiation of dendritic cell progenitors. J Immunol 1999; 162: 577–584
14 Antonysamy MA, Fanslow WC, Fu F et al. Evidence for a role of IL-17 in alloimmunity:
A novel IL-17 antagonist promotes heart graft survival. Transplant Proc 1999; 31: 93
15 Van Kooten C, Boonstra JG, Paape ME et al. Interleukin-17 activates human renal epi-
thelial cells in vitro and is expressed during renal allograft rejection. J Am Soc Nephrol 1998; 9: 1526–1534
16 Vanaudenaerde BM, Dupont LJ, Wuyts WA et al. The role of interleukin-17 during
acute rejection after lung transplantation. Eur Respir J 2006; 27: 779–787
17 Weaver CT, Hatton RD, Mangan PR, Harrington LE. IL-17 family cytokines and the
expanding diversity of effector T cell lineages. Annu Rev Immunol 2007; 25: 821–852 18 Miura M, El-Sawy T, Fairchild RL. Neutrophils mediate parenchymal tissue necrosis
and accelerate the rejection of complete major histocompatibility complex-disparate car-diac allografts in the absence of interferon-gamma. Am J Pathol 2003; 162: 509–519 19 Verma ND, Boyd R, Robinson C, Plain KM, Tran GT, Hall BM. Interleukin-12p70 pro-
longs allograft survival by induction of interferon gamma and nitric oxide production.
Transplantation 2006; 82: 1324–1333
20 Mangan PR, Harrington LE, O’Quinn DB et al. Transforming growth factor-beta
induces development of the T(H)17 lineage. Nature 2006; 441: 231–234
131
21 Kleinschek MA, Owyang AM, Joyce-Shaikh B et al. IL-25 regulates Th17 function in
autoimmune inflammation. J Exp Med 2007; 204: 161–170
22 Wood KJ, Sakaguchi S. Regulatory T cells in transplantation tolerance. Nat Rev Immu-
nol 2003; 3: 199–210
23 Kretschmer K, Apostolou I, Hawiger D, Khazaie K, Nussenzweig MC, von Boehmer H.
Inducing and expanding regulatory T cell populations by foreign antigen. Nat Immunol 2005; 6: 1219–1227
24 Bettelli E, Carrier Y, Gao W et al. Reciprocal developmental pathways for the generation
of pathogenic effector TH17 and regulatory T cells. Nature 2006; 441: 235–238
25 Veldhoen M, Hocking RJ, Atkins CJ, Locksley RM, Stockinger B. TG Fbeta in the
context of an inflammatory cytokine milieu supports de novo differentiation of IL-17-producing T cells. Immunity 2006; 24: 179–189
26 Xu L, Kitani A, Fuss I, Strober W. Cutting Edge: Regulatory T cells induce CD4+CD25–
Foxp3– T cells or are self-induced to become Th17 cells in the absence of exogenous TGF-beta. J Immunol 2007; 178: 6725–6729
27 Lohr J, Knoechel B, Wang JJ, Villarino A V, Abbas AK. Role of IL-17 and regulatory T
lymphocytes in a systemic autoimmune disease. J Exp Med 2006; 203: 2785–2791
28 Boros P, Bromberg JS. New cellular and molecular immune pathways in ischemia/reper-
fusion injury. Am J Transplant 2006; 6: 652–658
29 Perico N, Cattaneo D, Sayegh MH, Remuzzi G. Delayed graft function in kidney trans-
plantation. Lancet 2004; 364: 1814–1827
30 Lemay S, Rabb H, Postler G, Singh AK. Prominent and sustained up-regulation of
gp130-signaling cytokines and the chemokine MIP-2 in murine renal ischemia-reperfu-sion injury. Transplantation 2000; 69: 959–963
31 Kwon O, Molitoris BA, Pescovitz M, Kelly KJ. Urinary actin, interleukin-6, and inter-
leukin-8 may predict sustained ARF after ischemic injury in renal allografts. Am J Kid-ney Dis 2003; 41: 1074–1087
32 Patel NS, Chatterjee PK, Di Paola R et al. Endogenous interleukin-6 enhances the renal
injury, dysfunction, and inflammation caused by ischemia/reperfusion. J Pharmacol Exp Ther 2005; 312: 1170–1178
33 Kielar ML, John R, Bennett M et al. Maladaptive role of IL-6 in ischemic acute renal
failure. J Am Soc Nephrol 2005; 16: 3315–3325
34 Yoshida S, Haque A, Mizobuchi T et al. Anti-type V collagen lymphocytes that express
IL-17 and IL-23 induce rejection pathology in fresh and well-healed lung transplants.
Am J Transplant 2006; 6: 724–735
132
流式细胞仪检测技术与质量控制
流式细胞仪检测技术与质量 控制 -标准化文件发布号:(9456-EUATWK-MWUB-WUNN-INNUL-DDQTY-KII
流式细胞仪检测技术与质量控制 【摘要】流式细胞术(FCM)检测HLA等位基因是最近新建立的方法,与原有的分型技术相比,技术上有很大的改进和突破。流式细胞术已广泛用于临床常规检验中,为保证检验结果的可靠性,提高准确度和室间结果的可比性,流式细胞术质量控制越来越受到重视。它在功能水平上对单细胞或其他生物粒子进行定量分析和分选,同传统的荧光镜检查相比,具有速度快、精密度高、准确性好等特点。 【关键词】流式细胞术;检测技术;质量控制 流式细胞仪检验技术(FCM),即流式细胞术,是以流式细胞仪作为检测手段,以免疫荧光技术作为主要标记方法的一门先进的分析技术。该方法用免疫磁珠作为载体,在同一微孔内进行反应,利用流式细胞仪检测杂交信号和区分探针的种类。本技术使用的免疫磁珠具有一定的特性,磁珠可利用颜色进行标识[1]。当免疫磁珠上两种颜色混合的比例不同时,经流式细胞仪检测后即可区分定义为不同种类的免疫磁珠,目前两种颜色的组合在流式细胞仪上最多可区分成为100种不同的免疫磁珠。
1 材料与方法 1.1 标本收集 收集近3年本院治疗的30例患者,对30例患者行流式细胞仪检测,30例受检者中,男性患者16例,女性患者14例,最大年龄60岁,最小年龄17岁,患者平均年龄39岁。 2 检测方法 2.1 采用特定的免疫磁珠作为载体,将已知序列特异性探针(SSO)固定在免疫磁珠上,每一种特异性探针固定在已知颜色比例的免疫磁珠上。由于免疫磁珠上颜色比例的不同,在流式细胞仪红色激光束下可进行区分,根据事先设计的标记情况,通过流式细胞仪检测后可确认特定颜色比例免疫磁珠上携带的特异性探针的种类,从而达到将探针区分的目的。 2.2利用标记的特异性引物对目的DNA进行扩增,将PCR扩增产物与免疫磁珠上的序列特异性探针(SSO)在同一孔内进行特异性杂交,再加入荧光显色剂,然后利用流式细胞仪绿色激光束检测杂交信号,红色激光束区分探针的种类,利用软件分析杂交结果得出样本HLA基因型别。 3 方法学评价该方法与PCR-SSO有相似的地方,但是技术上有重大的突破。本方法灵敏度非常高,在
计算机原理试题与答案
全国2004年4月高等教育自学考试 计算机原理试题 课程代码:02384 第一部分选择题(共25分) 一、单项选择题(本大题共25小题,每小题1分,共25分) 在每小题列出的四个选项中只有一个选项是符合题目要求的,请将其代码填写在题后的括号内。错选、多选或未选均无分。 1.计算机中一次处理的最大二进制位数即为() A.位B.字节 C.字长D.代码 2.下列算式中属于逻辑运算的是() A.1+1=2 B.1-1=0 C.1+1=10 D.1+1=1 3.下图所示的门电路,它的逻辑表达式是() A.F=CD AB B.F=ABCD C.F=AB+CD D.F=ABCD 4.八进制数中的1位对应于二进制数的() A.2位B.3位 C.4位D.5位 5.下列叙述正确的是() A.原码是表示无符号数的编码方法 B.对一个数据的原码的各位取反而且在末位再加1就可以得到这个数据的补码
C.定点数表示的是整数 D.二进制数据表示在计算机中容易实现 6.浮点数0.00100011B×2-1的规格化表示是() A.0.1000110B×2-11B B.0.0100011B×2-10B C.0.0100011B×20B D.0.1000110B×21B 7.两个定点数作补码加法运算,对相加后最高位出现进位1的处理是() A.判为溢出B.AC中不保留 C.寄存在AC中D.循环加到末位 8.运算器中通用寄存器的长度一般取() A.8位B.16位 C.32位D.等于计算机字长 9.目前在大多数微型机上广泛使用宽度为32/64位的高速总线是() A.ISA B.EISA C.PCI D.VESA 10.某计算机指令的操作码有8个二进位,这种计算机的指令系统中的指令条数至多为 ()A.8 B.64 C.128 D.256 11.间接访内指令LDA @Ad的指令周期包含CPU周期至少有() A.一个B.二个 C.三个D.四个 12.在程序中,可用转移指令实现跳过后续的3条指令继续执行。这种指令的寻址方式是() A.变址寻址方式B.相对寻址方式
天线分集技术的原理
天线分集技术的原理 最初,许多设计者可能会担心区域规范的复杂性问题,因为在全世界范围内,不同区域规范也各异。然而,只要多加研究便能了解并符合不同区域的法规,因为在每一个地区,通常都会有一个政府单位负责颁布相关文件,以说明“符合特定目的的发射端相关的规则。 无线电通信中更难于理解的部分在于无线电通信链路质量与多种外部因素相关,多种可变因素交织在一起产生了复杂的传输环境,而这种传输环境通常很难解释清楚。然而,掌握基本概念往往有助于理解多变的无线电通信链接品质,一旦理解了这些基本概念,其中许多问题可以通过一种低成本、易实现的被称作天线分集(antenna diversity)的技术来实现。 环境因素的考虑 影响无线电通信链路持续稳定的首要环境因素是被称为多径/衰落和天线极化/分集的现象。这些现象对于链路质量的影响要么是建设性的要么是破坏性的,这取决于不同的特定环境。可能发生的情况太多了,于是,当我们试着要了解特定的环境条件在某个时间点对无线电通信链接的作用,以及会造成何种链接质量时,这无疑是非常困难的。 天线极化/分集 这种被称为天线极化的现象是由给定天线的方向属性引起的,虽然有时候把天线极化解释为在某些无线电通信链路质量上的衰减,但是一些无线电通信设计者经常利用这一特性来调整天线,通过限制收发信号在限定的方向范围之内达其所需。这是可行的,因为天线在各个方向上的辐射不均衡,并且利用这一特性能够屏蔽其他(方向)来源的射频噪声。 简单的说,天线分为全向和定向两种。全向天线收发信号时,在各个方向的强度相同,而定向天线的收发信号被限定在一个方向范围之内。若要打造高度稳固的链接,首先就要从了解此应用开始。例如:如果一个链路上的信号仅来自于特定的方向,那么选择定向天线获
计算机组成原理试题及答案
二、填空题 1 字符信息是符号数据,属于处理(非数值)领域的问题,国际上采用的字符系统是七单位的(ASCII)码。P23 2 按IEEE754标准,一个32位浮点数由符号位S(1位)、阶码E(8位)、尾数M(23位)三个域组成。其中阶码E的值等于指数的真值(e)加上一个固定的偏移值(127)。P17 3 双端口存储器和多模块交叉存储器属于并行存储器结构,其中前者采用(空间)并行技术,后者采用(时间)并行技术。P86 4 衡量总线性能的重要指标是(总线带宽),它定义为总线本身所能达到的最高传输速率,单位是(MB/s)。P185 5 在计算机术语中,将ALU控制器和()存储器合在一起称为()。 6 数的真值变成机器码可采用原码表示法,反码表示法,(补码)表示法,(移码)表示法。P19-P21 7 广泛使用的(SRAM)和(DRAM)都是半导体随机读写存储器。前者的速度比后者快,但集成度不如后者高。P67 8 反映主存速度指标的三个术语是存取时间、(存储周期)和(存储器带宽)。P67 9 形成指令地址的方法称为指令寻址,通常是(顺序)寻址,遇到转移指令时(跳跃)寻址。P112 10 CPU从(主存中)取出一条指令并执行这条指令的时间和称为(指令周期)。 11 定点32位字长的字,采用2的补码形式表示时,一个字所能表示
的整数范围是(-2的31次方到2的31次方减1 )。P20 12 IEEE754标准规定的64位浮点数格式中,符号位为1位,阶码为11位,尾数为52位,则它能表示的最大规格化正数为(+[1+(1-2 )]×2 )。 13 浮点加、减法运算的步骤是(0操作处理)、(比较阶码大小并完成对阶)、(尾数进行加或减运算)、(结果规格化并进行舍入处理)、(溢出处理)。P54 14 某计算机字长32位,其存储容量为64MB,若按字编址,它的存储系统的地址线至少需要(14)条。64×1024KB=2048KB(寻址范32围)=2048×8(化为字的形式)=214 15一个组相联映射的Cache,有128块,每组4块,主存共有16384块,每块64个字,则主存地址共(20)位,其中主存字块标记应为(9)位,组地址应为(5)位,Cache地址共(13)位。 16 CPU存取出一条指令并执行该指令的时间叫(指令周期),它通常包含若干个(CPU周期),而后者又包含若干个(时钟周期)。P131 17 计算机系统的层次结构从下至上可分为五级,即微程序设计级(或逻辑电路级)、一般机器级、操作系统级、(汇编语言)级、(高级语言)级。P13 18十进制数在计算机内有两种表示形式:(字符串)形式和(压缩的十进制数串)形式。前者主要用在非数值计算的应用领域,后者用于直接完成十进制数的算术运算。P19 19一个定点数由符号位和数值域两部分组成。按小数点位置不同,
触摸屏的种类及工作原理
触摸屏种类及原理 随着多媒体信息查询的与日俱增,人们越来越多地谈到触摸屏,因为触摸屏不仅适用于中国多媒体信息查询的国情,而且触摸屏具有坚固耐用、反应速度快、节省空间、易于交流等许多优点。利用这种技术,我们用户只要用手指轻轻地碰计算机显示屏上的图符或文字就能实现对主机操作,从而使人机交互更为直截了当,这种技术大大方便了那些不懂电脑操作的用户。 触摸屏作为一种最新的电脑输入设备,它是目前最简单、方便、自然的一种人机交互方式。它赋予了多媒体以崭新的面貌,是极富吸引力的全新多媒体交互设备。触摸屏在我国的应用范围非常广阔,主要是公共信息的查询;如电信局、税务局、银行、电力等部门的业务查询;城市街头的信息查询;此外应用于领导办公、工业控制、军事指挥、电子游戏、点歌点菜、多媒体教学、房地产预售等。将来,触摸屏还要走入家庭。 随着使用电脑作为信息来源的与日俱增,触摸屏以其易于使用、坚固耐用、反应速度快、节省空间等优点,使得系统设计师们越来越多的感到使用触摸屏的确具有具有相当大的优越性。触摸屏出现在中国市场上至今只有短短的几年时间,这个新的多媒体设备还没有为许多人接触和了解,包括一些正打算使用触摸屏的系统设计师,还都把触摸屏当作可有可无的设备,从发达国家触摸屏的普及历程和我国多媒体信息业正处在的阶段来看,这种观念还具有一定的普遍性。事实上,触摸屏是一个使多媒体信息或控制改头换面的设备,它赋予多媒体系统以崭新的面貌,是极富吸引力的全新多媒体交互设备。发达国家的系统设计师们和我国率先使用触摸屏的系统设计师们已经清楚的知道,触摸屏对于各种应用领域的电脑已经不再是可有可无的东西,而是必不可少的设备。它极大的简化了计算机的使用,即使是对计算机一无所知的人,也照样能够信手拈来,使计算机展现出更大的魅力。解决了公共信息市场上计算机所无法解决的问题。 随着城市向信息化方向发展和电脑网络在国民生活中的渗透,信息查询都已用触摸屏实现--显示内容可触摸的形式出现。为了帮助大家对触摸屏有一个大概的了解,笔者就在这里提供一些有关触摸屏的相关知识,希望这些内容能对大家有所用处。 一、触摸屏的工作原理 为了操作上的方便,人们用触摸屏来代替鼠标或键盘。工作时,我们必须首先用手指或其它物体触摸安装在显示器前端的触摸屏,然后系统根据手指触摸的图标或菜单位置来定位选择信息输入。触摸屏由触摸检测部件和触摸屏控制器组成;触摸检测部件安装在显示器屏幕前面,用于检测用户触摸位置,接受后送触摸屏控制器;而触摸屏控制器的主要作用是从触摸点检测装置上接收触摸信息,并将它转换成触点坐标,再送给CPU,它同时能接收CPU发来的命令并加以执行。 二、触摸屏的主要类型
BD分析TH17细胞
流式细胞术分析TH17细胞 1.肝素钠真空抗凝采血管取人外周血全血1-2ml,血液室温放置,8小时内进行检测,否则需弃用。 2.取5个流式管,分别标记空白管、CD4抗体单染管、IL-17A抗体单染管、IL-17A同型对照抗体管、待测管,每个流式管分别加入100μl全血,用RPMI1640(不含FBS)1:1等体积稀释。 3.刺激细胞活化:向稀释后的全血样本管加入2μl BD Leukocyte Activation Cocktail with BD GolgiPlug,货号550583(刺激剂和蛋白转运抑制剂),混匀。37o C,5%CO2培养箱或37o C水浴孵育4-6小时(不超过6个小时)。 4.细胞表面标记染色:向CD4抗体单染管、IL-17A同型对照抗体管、待测管这三管分别加入1Test体积的CD4抗体,室温避光孵育15-30分钟。 5.制备红细胞裂解液:制备1X.红细胞裂解液(BD Lysing Buffer10X,货号555899),取10X红细胞裂解液,按照体积.1(10X红细胞裂解液):9(dH2O),稀释。 6.每100μl全血加入2ml1X红细胞裂解液,室温裂解15-30min,至细胞悬液呈澄清透明状。 7.裂红后,500g,离心5分钟,洗涤细胞,离心后弃上清。 8.加入1ml FBS(货号554656),500g,离心5分钟,洗涤细胞,离心后弃上清。 9.破膜固定液制备(货号554714),554714包括Fixation/Permeabilization solution和BD Perm/Wash?Buffer(10X)。Perm/Wash?Buffer(10X)需要稀释:1体积BD Perm/Wash?Buffer(10X),9体积distilled H20,1:9混合,配制成1XPerm/Wash?Buffer 10.细胞固定:涡旋细胞3秒钟,每管加入250μL Fixation/Permeabilization solution,涡旋混匀,4°C孵育20分钟。 11.破膜:每管直接加入1ml1X Perm/Wash Working Solution到固定的细胞中,4°C,500g离心10分钟,弃上清。 12.每管加入1ml1X Perm/Wash Working Solution,4°C500g离心10分钟,弃上清。
计算机组成原理参考答案汇总
红色标记为找到了的参考答案,问答题比较全,绿色标记为个人做的,仅供参考!第一章计算机系统概述 1. 目前的计算机中,代码形式是______。 A.指令以二进制形式存放,数据以十进制形式存放 B.指令以十进制形式存放,数据以二进制形式存放 C.指令和数据都以二进制形式存放 D.指令和数据都以十进制形式存放 2. 完整的计算机系统应包括______。 A. 运算器、存储器、控制器 B. 外部设备和主机 C. 主机和实用程序 D. 配套的硬件设备和软件系统 3. 目前我们所说的个人台式商用机属于______。 A.巨型机 B.中型机 C.小型机 D.微型机 4. Intel80486是32位微处理器,Pentium是______位微处理器。 A.16B.32C.48D.64 5. 下列______属于应用软件。 A. 操作系统 B. 编译系统 C. 连接程序 D.文本处理 6. 目前的计算机,从原理上讲______。 A.指令以二进制形式存放,数据以十进制形式存放 B.指令以十进制形式存放,数据以二进制形式存放 C.指令和数据都以二进制形式存放 D.指令和数据都以十进制形式存放 7. 计算机问世至今,新型机器不断推陈出新,不管怎样更新,依然保有“存储程序”的概念,最早提出这种概念的是______。 A.巴贝奇 B.冯. 诺依曼 C.帕斯卡 D.贝尔 8.通常划分计算机发展时代是以()为标准 A.所用的电子器件 B.运算速度 C.计算机结构 D.所有语言 9.到目前为止,计算机中所有的信息任以二进制方式表示的理由是() A.节约原件 B.运算速度快 C.由物理器件的性能决定 D.信息处理方便 10.冯.诺依曼计算机中指令和数据均以二进制形式存放在存储器中,CPU区分它们的依据是() A.指令操作码的译码结果 B.指令和数据的寻址方式 C.指令周期的不同阶段 D.指令和数据所在的存储单元 11.计算机系统层次结构通常分为微程序机器层、机器语言层、操作系统层、汇编语言机器层和高级语言机器层。层次之间的依存关系为() A.上下层都无关 B.上一层实现对下一层的功能扩展,而下一层与上一层无关 C.上一层实现对下一层的功能扩展,而下一层是实现上一层的基础
浅析发射分集与接收分集技术
浅析发射分集与接收分集技术 1 概述 1.1 多天线信息论简介 近年来,多天线系统(也称为MIMO系统)引起了人们很大的研究兴趣,多天线系统原理如图1所示,它可以增加系统的容量,改进误比特率(BER).然而,获得这些增益的代价是硬件的复杂度提高,无线系统前端复杂度、体积和价格随着天线数目的增加而增加。使用天线选择技术,就可以在获得MIMO系统优势的同时降低成本。 图1 MIMO系统原理 有两种改进无线通信的方法:分集方法、复用方法。分集方法可以提高通信系统的鲁棒性,利用发送和接收天线之间的多条路径,改善系统的BER。在接收端,这种分集与RAKE接收提供的类似。分集也可以通过使用多根发射天线来得到,但是必须面对发送时带来的相互干扰。这一类主要是空时编码技术。 另外一类MIMO技术是空间复用,来自于这样一个事实:在一个具有丰富散射的环境中,接收机可以解析同时从多根天线发送的信号,因此,可以发送并行独立的数据流,使得总的系统容量随着min( , )线性增长,其中
和 是接收和发送天线的数目。 1.2 空时处理技术 空时处理始终是通信理论界的一个活跃领域。在早期研究中,学者们主要注重空间信号传播特性和信号处理,对空间处理的信息论本质探讨不多。上世纪九十年代中期,由于移动通信爆炸式发展,对于无线链路传输速率提出了越来越高的要求,传统的时频域信号设计很难满足这些需求。工业界的实际需求推动了理论界的深入探索。 在MIMO技术的发展,可以将空时编码的研究分为三大方向:空间复用、空间分集与空时预编码技术,如图2所示。 图2 MIMO技术的发展
1.3 空间分集研究 多天线分集接收是抗衰落的传统技术手段,但对于多天线发送分集,长久以来学术界并没有统一认识。1995年Telatarp[3]首先得到了高斯信道下多天线发送系统的信道容量和差错指数函数。他假定各个通道之间的衰落是相互独立的。几乎同时, Foschini和Gans在[4]得到了在准静态衰落信道条件下的截止信道容量(Outage Capacity)。此处的准静态是指信道衰落在一个长周期内保持不变,而周期之间的衰落相互独立,也称这种信道为块衰落信道(Block Fading)。 Foschini和Gans的工作,以及Telatar的工作是多天线信息论研究的开创 性文献。在这些著作中,他们指出,在一定条件下,采用多个天线发送、多个天线接收(MIMO)系统可以成倍提高系统容量,信道容量的增长与天线数目成线性关系 1.4 空时块编码 (STBC) 本文我们主要介绍一类高性能的空时编码方法——空时块编码( STBC: Space Time Block Code)。 STBC编码最先是由Alamouti[1]在1998年引入的,采用了简单的两天线发分集编码的方式。这种STBC编码最大的优势在于,采用简单的最大似然译码准则,可以获得完全的天线增益。 Tarokh[5]进一步将2天线STBC编码推广到多天线形式,提出了通用的正交设计准则。 2 MIMO原理及方案
快速体外分离、长期培养人Th17细胞
快速体外分离、长期培养人Th17细胞方法 1.Subject 全血 2.CD4+T细胞浓缩 采用RosetteSep(StemCell Technologies,Canada)进行阴性选择得到浓缩CD4+T细胞。建立标准密度梯度离心,分离出淋巴细胞 3.CD4+T细胞刺激 将浓缩的CD4+T细胞悬浮在RPMI1640培养基中(Sigma),包含青霉素/链霉素,L-谷氨酰胺,HEPES缓冲液和10%小牛血清(R10),浓度为1×106个细胞/ ml.含 3×106个细胞的3mlR10,在15ml Falcon管中,用每毫升含4μL(1mg/ml)PMA和2μL(1mg/ml)伊沃诺霉素(AG Scientific,San Diego,CA)的细胞溶液刺激。继而加入总量3μL的包含CD49d (1mg/ml)和CD28(0.5mg/ml)的共刺激抗体(BD Biosciences)。随后细胞在37℃、5%CO2条件下孵化3个半小时。阴性对照采用相似方法,但用R10替换PMA和伊沃诺霉素。为了用细胞内细胞因子标记法测量IL-17产量,浓缩的CD4+T细胞用10μL Brefeldin A(1mg/ml)刺激。 4.IL-17捕捉复合物的准备 IL-17捕捉复合物由2个生物素标记的抗体组成,2个抗体通过一个抗生物素蛋白分子连接。直接连在T细胞表面CD45分子的抗体与IL-17抗体配对。2μL生物素标记的CD45抗体(clone H130)(Caltag,Invirtrogen,CA)和20μL生物素标记的IL-17抗体(0.5mg/ml)(clone:eBio64DEC17)(ebiosciences,CA)混合加入eppendorf管,并彻底漩涡。然后,加入2μL未作标记的5mg/ml的生物素(Invitrogen,CA),并立即充分混匀。复合物在室温下孵化10分钟,使用前再次漩涡。 5.IL-17体外捕获分析 被刺激过的细胞用含有2%小牛血清(2%FCS/PBS)的冰冻PBS(Cellgro ,Mediatech,V A),冲洗,以500×克离心10分钟成小球,上清液被完全吸出。然后细胞被悬浮在100μL2%的FCS/PBS,将20μL捕获的IL-17复合体加入。冰上孵化15分钟后,加入9mlR10。将试管放入转动器,在37℃和5%CO2条件下额外孵化1个半小时。 6.IL-17+细胞分选 被捕获的IL-17细胞可以用2种技术分选:一种是FACS Aria细胞分选器(BD),另一种是磁珠技术(Miltenyi,Germany)。FACS Aria细胞分选器,细胞被一种太平洋蓝活性胺标记来区分活的和死的细胞。然后用FACS/PBS冲洗细胞,细胞表面用10μL的CD3-FITC,5μL的CD4-Alexa700(BD BIOsciences)和20μL(0.25μg)IL-17A-PE(clone:eBio64CAP17)抗体标记。细胞在暗室诗文条件下孵化15分钟,分析前用2%FCS/PBS冲洗一次。 磁珠法捕获的细胞用IL-17A-PE标记15分钟。用2%FCS/PBS冲洗一次将多余的抗体移除,然后参看说明将细胞在4℃条件下加抗-PE磁珠孵化。简而言之,再次冲洗细胞并使之悬浮在500μL缓冲液中。悬浮液用洗液管加入MS柱(Miltenyi,German)并放到磁体上,用500μL的PBS洗3次。将虑柱从磁体上拿下,被选择的细胞收集在一个15mL的Falcon管。被选择的细胞再次通过磁性分离进一步浓缩。 Aria分选或磁珠净化得到的Th17细胞,培养在包含50单位的人IL-2(NIH/Roche,switzerland)(R10-50)的R10培养基,. 2天后,细胞被250.000照射feeders(3000 rad)和抗-CD3/抗-CD8双特异性单克隆抗体(1μg)刺激。每3周细胞被照射feeders(3000 rad)
流式细胞仪检测技术与质量控制
流式细胞仪检测技术与质量控制 【摘要】流式细胞术(FCM)检测HLA等位基因是最近新建立的方法,与原有的分型技术相比,技术上有很大的改进和突破。流式细胞术已广泛用于临床常规检验中,为保证检验结果的可靠性,提高准确度和室间结果的可比性,流式细胞术质量控制越来越受到重视。它在功能水平上对单细胞或其他生物粒子进行定量分析和分选,同传统的荧光镜检查相比,具有速度快、精密度高、准确性好等特点。 【关键词】流式细胞术;检测技术;质量控制 流式细胞仪检验技术(FCM),即流式细胞术,是以流式细胞仪作为检测手段,以免疫荧光技术作为主要标记方法的一门先进的分析技术。该方法用免疫磁珠作为载体,在同一微孔内进行反应,利用流式细胞仪检测杂交信号和区分探针的种类。本技术使用的免疫磁珠具有一定的特性,磁珠可利用颜色进行标识[1]。当免疫磁珠上两种颜色混合的比例不同时,经流式细胞仪检测后即可区分定义为不同种类的免疫磁珠,目前两种颜色的组合在流式细胞仪上最多可区分成为100种不同的免疫磁珠。
1 材料与方法 1.1 标本收集 收集近3年本院治疗的30例患者,对30例患者行流式细胞仪检测,30例受检者中,男性患者16例,女性患者14例,最大年龄60岁,最小年龄17岁,患者平均年龄39岁。 2 检测方法 2.1 采用特定的免疫磁珠作为载体,将已知序列特异性探针(SSO)固定在免疫磁珠上,每一种特异性探针固定在已知颜色比例的免疫磁珠上。由于免疫磁珠上颜色比例的不同,在流式细胞仪红色激光束下可进行区分,根据事先设计的标记情况,通过流式细胞仪检测后可确认特定颜色比例免疫磁珠上携带的特异性探针的种类,从而达到将探针区分的目的。 2.2利用标记的特异性引物对目的DNA进行扩增,将PCR扩增产物与免疫磁珠上的序列特异性探针(SSO)在同一孔内进行特异性杂交,再加入荧光显色剂,然后利用流式细胞仪绿色激光束检测杂交信号,红色激光束区分探针的种类,利用软件分析杂交结果得出样本HLA基因型别。 3 方法学评价该方法与PCR-SSO有相似的地方,但是技术上有重大的突破。本方法灵敏度非常高,
计算机组成原理试题及答案
《计算机组成原理》试题 一、(共30分) 1.(10分) (1)将十进制数+107/128化成二进制数、八进制数和十六进制数(3分) (2)请回答什么是二--十进制编码?什么是有权码、什么是无权码、各举一个你熟悉的有权码和无权码的例子?(7分) 2.已知X=0.1101,Y=-0.0101,用原码一位乘法计算X*Y=?要求写出计算过程。(10分) 3.说明海明码能实现检错纠错的基本原理?为什么能发现并改正一位错、也能发现二位错,校验位和数据位在位数上应满足什么条件?(5分) 4.举例说明运算器中的ALU通常可以提供的至少5种运算功能?运算器中使用多累加器的好处是什么?乘商寄存器的基本功能是什么?(5分) 二、(共30分) 1.在设计指令系统时,通常应从哪4个方面考虑?(每个2分,共8分) 2.简要说明减法指令SUB R3,R2和子程序调用指令的执行步骤(每个4分,共8分) 3.在微程序的控制器中,通常有哪5种得到下一条指令地址的方式。(第个2分,共10分) 4.简要地说明组合逻辑控制器应由哪几个功能部件组成?(4分) 三、(共22分) 1.静态存储器和动态存储器器件的特性有哪些主要区别?各自主要应用在什么地方?(7分) 2.CACHE有哪3种基本映象方式,各自的主要特点是什么?衡量高速缓冲存储器(CACHE)性能的最重要的指标是什么?(10分) 3.使用阵列磁盘的目的是什么?阵列磁盘中的RAID0、RAID1、RAID4、RAID5各有什么样的容错能力?(5分) 四、(共18分) 1.比较程序控制方式、程序中断方式、直接存储器访问方式,在完成输入/输出操作时的优缺点。(9分) 2.比较针式、喷墨式、激光3类打印机各自的优缺点和主要应用场所。(9分) 答案 一、(共30分) 1.(10分) (1) (+107/128)10 = (+1101011/10000000)2 = (+0.1101011)2 = (+0.153)8 = (+6B)16 (2) 二-十进制码即8421码,即4个基2码位的权从高到低分别为8、4、2、1,使用基码的0000,0001,0010,……,1001这十种组合分别表示0至9这十个值。4位基二码之间满足二进制的规则,而十进制数位之间则满足十进制规则。 1
喷码机触摸屏的工作原理与应用
喷码机触摸屏的工作原理与应用 一、触摸屏的工作原理为了操作上的方便,人们用触摸屏来代替鼠标或键盘。工作时,我们必须首先用手指或其它物体触摸安装在显示器前端的触摸屏,然后系统根据手指触摸的图标或菜单位置来定位选择信息输入。触摸屏由触摸检测部件和触摸屏控制器组成;触摸检测部件安装在显示器屏幕前面,用于检测用户触摸位置,接受后送触摸屏控制器;而触摸屏控制器的主要作用是从触摸点检测装置上接收触摸信息,并将它转换成触点坐标,再送给CPU,它同时能接收CPU 发来的命令并加以执行。二、触摸屏的主要类型从技术原理来区别触摸屏,可分为五个基本种类:矢量压力传感技术触摸屏、电阻技术触摸屏、电容技术触摸屏、红外线技术触摸屏、表面声波技术触摸屏。其中矢量压力传感技术触摸屏已退出历史舞台。触摸屏红外屏价格低廉,但其外框易碎,容易产生光干扰,曲面情况下失真;电容屏设计理论好,但其图象失真问题很难得到根本解决;电阻屏的定位准确,但其价格颇高,且怕刮易损。表面声波触摸屏解决了以往触摸屏的各种缺陷,清晰抗暴,适于各种场合,缺憾是屏表面的水滴、尘土会使触摸屏变的迟钝,甚至不工作。按照触摸屏的工作原理和传输信息的介质,我们把触摸屏分为四种,它们分别为电阻式、红外线式、电容感应式以及表面声波式,下面笔者就对上述的各种类型的触摸屏进行简要介绍: 1、电阻式触摸屏电阻触摸屏的屏体部分是一块与显示器表面非常配合的多层复合薄膜,由一层玻璃或有机玻璃作为基层,表面涂有一层透明的导电层(OTI,氧化铟),上面再盖有一层外表面硬化处理、光滑防刮的塑料层,它的内表面也涂有一层OTI,在两层导电层之间有许多细小(小于千分之一英寸)的透明隔离点把它们隔开绝缘。当手指接触屏幕,两层OTI 导电层出现一个接触点,因其中一面导电层接通Y轴方向的5V 均匀电压场,使得侦测层的电压由零变为非零,控制器侦测到这个接通后,进行A/D 转换,并将得到的电压值与5V 相比,即可得触摸点的Y 轴坐标,同理得出X 轴的坐标,这就是电阻技术触摸屏共同的最基本原理。电阻屏根据引出线数多少,分为四线、五线等多线电阻触摸屏。五线电阻触摸屏的A面是导电玻璃而不是导电涂覆层,导电玻璃的工艺使其的寿命得到极大的提高,并且可以提高透光率。 电阻式触摸屏的OTI 涂层比较薄且容易脆断,涂得太厚又会降低透光且形成内反射降低清晰度,OTI 外虽多加了一层薄塑料保护层,但依然容易被锐利物件所破坏;且由于经常被触动,表层OTI 使用一定时间后会出现细小裂纹,甚至变型,如其中一点的外层OTI 受破坏而断裂,便失去作为导电体的作用,触摸屏的寿命并不长久。但电阻式触摸屏不受尘埃、水、污物影响。这种触摸屏利用压力感应进行控制。它用两层高透明的导电层组成触摸屏,两层之间距离仅为2.5 微米。当手指按在触摸屏上时,该处两层导电层接触,电阻发生变化,在X 和Y 两个方向上产生信号,然后送触摸屏控制器。这种触摸屏能在恶劣环境下工作,但手感和透光性较差,适合配带手套和不能用手直接触控的场合。电阻类触摸屏的关键在于材料科技,常用的透明导电涂层材料有:A、ITO,氧化铟,弱导电体,特性是当厚度降到1800 个(埃=10-10 米)以下时会突然变得透明,透光率为80%,再薄下去透光率反而下降,到300 埃厚度时又上升到80%。ITO 是所有电阻技术触摸屏及电容技术触摸屏都用到的主要材料,实际上电阻和电容技术触摸屏的工作面就是ITO 涂层。B、镍金涂层,五线电阻触摸屏的外层导电层使用的是延展性好的镍金涂层材料,外导电层由于频繁触摸,使用延展性好的镍金材料目的是为了延长使用寿命,但是工艺成本较为高昂。镍金导电层虽然延展性好,但是只能作透明导体,不适合作为电阻触摸屏的工作面,因为它导电率高,而且金属不易做到厚度非常均匀,不宜作电压分布层,只能作为探层。 2、电容式触摸屏电容式触摸屏的构造主要是在玻璃屏幕上镀一层透明的薄膜体层,再在导体层外加上一块保护玻璃,双玻璃设计能彻底保护导体层及感应器。电容式触摸屏在
新型的CD_4_T细胞_Th17细胞
[8]Grunds tro m S,And ers on P,Scheipers P,et a l.Bcl23and NFkap2 paB p502p50homod i m ers act as transcri pti onal rep ress ors i n tol2 erant CD4+T cells[J].J Bi ol Ch e m,2004,279(9): 846028468. [9]Y e S,W atts GF,M anda li a S,et al.Preli m i nary report:geneti c vari2 ati on i n t he hum an s tro m el ysi n pro m oter is associat ed w i th p rogers2 si on of coronary at heroscl erosis[J].BrH eart J,1995,73(3):2092 215. [10]H i rash i k iA,Ya m ada Y,M urase Y,et a l.Associ ati on of gen e po l y2 m orphis m s w i th coronary artery d is ease i n lo w2or h i gh2ris k s ub2 jects defined by conven ti onal ri sk factors[J].J Am Coll Cardio,l 2003,42(8):142921437. [11]Zh ou X,H uang J,Chen J,et al.H ap l otype anal ys i s of t he m atrix m etall op rotei nas e23gene and m yocard i al i n farcti on i n a Ch i n ese H an popu l ati on.Th e Beiji ng A t eroscl erosis Study[J].Thro m b H ae most,2004,92(4):8672873. [12]Poll anen PJ,Leh ti m ak iT,M i kkelsson J,et al.M atri xm etall oprotei2 nase3and9gene pro m oter pol ymorph is m s:j oi n t acti on of t w o l oci as a ris k factor for coronary artery co m p li cated p l aques[J].A t h er2 oscl erosis,2005,180(1):73278. [13]H oppm ann P,K och W,Schom i g A,et al.The5A/6A pol ymor2 ph is m of the stro m el ys i n21gene and res t en os i s after percu t an eou s coronary i n terventi on s[J].Eur H eart J,2004,25(4):3352341. [14]Y e S,Gale C R,M artyn C N.Vari ation i n t he m atri x m etalloprotei n2 ase21gen e and ris k of coronary heart disease[J].Eu r H eart J, 2003,24(18):166821671. [15]H l at ky MA,Ash l ey E,Querter m ou s T,et al.M atrix m etalloprotei n2 ase circu l ati ng l evels,gen eti c pol y m orph i s m s,and suscep ti b ili ty to acu t e m yocard i al i nfarcti on a m ong patients w it h coronary artery d isease[J].Am H eart J,2007,154(6):104321051.[16]Y a m ada Y,Iza wa H,Ic h i h ara S,et a l.Pred icti on of t h e ri sk ofm y2 ocard i al infarcti on from pol y m orph i s m s i n cand i date genes[J].N Engl J M ed,2002,347(24):191621923. [17]Samnegard A,Sil vei ra A,Lundman P,et a l.Seru m m atri x m et a ll o2 p rotei nas e23con centrati on i s i n fluenced by MMP2316125A/6A pro m oter genot ype and as soci ated wit h m yocard i al i n farcti on[J].J IntM ed,2005,258(5):4112419. [18]Gn asso A,Motti C,Irace C,et al.G enetic vari ati on i n hum an stro m elys i n gen e p ro m ot er and co m m on carotid geo m etry in healthy m ale sub jects[J].A rteri oscl er Th ro m b Vasc Bi o,l2000,20(6): 160021605. [19]Raura maa R,Va i sanen SB,Luong LA,et a l.Stro m el ys i n21and i n2 terleuk i n26gen e pro m oter po l y m orph is m s are deter m i nan ts of a2 s ymp t o m ati c caroti d artery at h eroscl erosis[J].A rteri oscler Thromb Vasc Bi o,l2000,20(12):265722662. [20]Rundek T,E l k i nd MS,P itt m an J,et al.Caroti d i nti m a2m ed i a thickness i s associ ated wit h all elic vari an ts of s tro m el ysi n21,i n ter2 leuk i n26,and h epatic li pase genes:t h e Northern Manh attan Pro2 s pecti ve Cohort Study[J].Stroke,2002,33(5):142021423. [21]Gh ilardiG,Bi ond iML,D e M on tiM,et al.M atrixm etall op rotei nas e2 1and m atri x m etall op rotei nas e23gen e p ro m ot er pol y m orph i s m s are associat ed w it h caroti d artery stenosis[J].Stroke,2002,33(10): 240822412. [22]A r m strong C,Ab illei ra S,Si tzerM,et al.Po l y m orph is m s i n MM P Fa m il y and TI M P genes and caroti d art ery i n ti m a2m ed ia th i ckness [J].Stroke,2007,38(11):289522899. [23]F l ex A,Gaetan i E,Papal eo P,et al.Proi n fla m m atory geneti c pro2 fil es i n sub jects w it h h i story of i sche m i c strok e[J].Stroke,2004, 35(10):227022275. 收稿日期:2008203217修回日期:2008205207 新型的CD4+T细胞:Th17细胞 王国栋,张爱元,邵静波 (潍坊市人民医院心内一科,山东潍坊261041) 中图分类号:R392.11文献标识码:A文章编号:100622084(2008)1722603204 摘要:Th17是近年来发现的一种CD4+T细胞亚型,在自身免疫性疾病中发挥重要的作用,其分泌产生几种致炎细胞因子,包括新发现的细胞因子白细胞介素17。Th17产生的细胞因子与Th1、Th2不同并且与其相互对抗。Th17细胞很可能对防御胞外病原菌的感染及自身免疫性疾病产生影响。影响Th17细胞从T细胞前体分化的因素也很快被发现,本文综述了Th17细胞诱导分化的影响因素、产生的细胞因子、在自身免疫病中的作用。 关键词:C D 4 +T细胞;Th17细胞;白细胞介素17 A N e w Sub s et of CD4+T C ell s:Th17WA NG Gu o2d ong,Z HA NG Ai2yuan,S HAO J i ng2bo.(De part2 m e n t of Card iol ogy,Wei fang P eo p le c s H os pit a l,Wei fang261041,China) Ab stract:Th17,a novel s ubset of CD4+T cells was i den ti fi ed t hat p l ays a cruci a l ro l e i n i n fla m2 m ation and au t oi m m un e d iseas e,wh ich p roduces s everal proi n fl a mm at ory cytok i n es,i ncl ud i ng t he novel cytok i ne i n t erl euk i n I L217.The cytok i nes produ ced by Th17are d ifferent fro m t hose by Th1or Th2, they are antagon i zed.Th17cells are prob ab l y eff ecti ve i n p rotection agai ns t extracell u lar b acteria,bu t al so p l ay a rol e i n a m p lification of au t o i m m une d i sorders.And t h e factors t hat s pecif y d ifferenti ati on of I L217producing effect or T cells fro m n ai ve T cell p recu rsors are be i ng rapidly d i scovered.Th is articl e discussed t h e fact ors on Th17i ndu ced generati on and d ifferenti ation,t he cytok i ne profili ng,the f un cti on of the cell li neage involved i n au t o i m m une d i seases. K ey word s:CD4+T cel;l Th17cel;l In t erleuk i n217 天然CD4+T细胞是一种具有很多亚群的前体,具 有抗原识别特异性,是主要组织相容性复合物ò类分子限制性T细胞。根据产生淋巴因子的种类和生物学功能的不同,CD4+T细胞主要分为辅助性T细胞1型(T help ce ll1, Th1)、2型(T help ce ll2,Th2)和调节性T细胞(regulatory T cells,T reg)[1]。Th1型细胞主要产生干扰素C(interferon2C, I FN2C)、白细胞介素2(i n terleu2 k i n22,I L22)和肿瘤坏死因子B (tumor necr osis f actor B,T NF2B),参与细胞免疫和迟发型超敏反应,协助B细胞产生Ig G2,在抗 细胞内微生物感染和细胞免疫所引起的炎性反应中发挥作用。Th2型细胞主要产生I L24、I L25、I L210和