电子版-生物膜动力学的研究现状与展望

生物膜动力学的研究现状与展望
1 引言
生物膜法作为一种高效的废水处理方法,已经在工业界获得了广泛应用。生物膜废水处理系统的性能在很大程度上取决于生物膜的形成及其动力学过程。最近三十年来,各国学者围绕生物膜的形成、发展、结构以及动力学特性等从数学模型、数值模拟和实验研究等方面进行了大量的研究,取得了许多重要进展,为生物膜反应器的设计提供了理论和实验支持,有力地推动了生物膜废水处理工艺的发展。
2 生物膜动力学模型的研究进展
动力学数学模型一直被作为模拟生物膜中微生物动力学行为和生物膜微观结构的一种有力工具,也是将生物膜内微观现象和大规模工艺运行的宏观指标联系起来的关键工具【1】。迄今为止,生物膜动力学数学模型的使用仍在研究领域占主导地位。科研工作者对生物膜形成、构成、结构及功能的兴趣,极大地推动了生物膜动力学数学模型的发展。自20世纪70年代反应-扩散动力学模型提出以来,描述生物膜动力学的模型先后又有Capdeville 增长动力学体系、元胞自动机模型和复合生物膜模型,分别介绍如下:
2.1 反应-扩散动力学模型【2,3】
反应-扩散动力学模型是描述生物膜动力学的最基本的模型。几乎所有的生物膜数学模型都假定生物膜内电子供体、电子受体和所有的营养物质只通过扩散作用传递给微生物(内部传质),而忽略了这些物质从液相主体到生物膜的传递过程(外部传质)。反应-扩散模型将生物膜假设为规则连续介质的稳态膜(包含单一物种),仅考虑一维(1D)物质传输和生化转化作用。生物膜被理想化成具有恒定厚度(f L )和统一细胞密度(f X )的薄膜。从液相主体到生物膜的基质通量是由生物膜内部的微生物活性产生。微生物增长用Monod 方程表示;基质消耗速率(ut r )假定正比于微生物生长速率;基质通量仅用扩散表示。生物膜外部传质限制被认为出现在位于生物膜和液相主体交界面处具有恒定厚度(f L )的边界层中。传质通量采用菲克定律(Fick Law)描述,但其中的扩散系数用有效扩散系数替代:S S e dS J D dx
=。这种理想化生物膜的数学模型可以用如下微分方程来表示22?.s S S e f S S
S d S q S D X t dx K S ?=-?+,0f x L ≤≤(1)
边界条件为0x =时0S dS dx =(2)f x L =时()S S S e L Sb S dS J D k S S dx ==-(3) 基质利用和扩散由方程(1)描述,边界条件采用式(2)和(3)描述。由于附着表面不可穿透,故此处的通量和基质梯度为零(见式(2))。在生物膜和液相主体交界面处的基质浓度(s S )由质量守恒式确定。即,通过边界层的基质通量必定等于进入生物膜的基质通量(见式(3))。这个理想化的数学模型可以利用有限差分法近似求解。当生物膜处于稳态时,系统可以使用有效因子法和伪解析法求解。关于有效因子法和伪解析法的详细介绍可以参考文献【2,3】。 生物膜反应-扩散理论自20世纪70年代提出后,经过各国学者的大量研究工作而得到完善,并得到了广泛接受和承认。然而,最近十几年来,许多新的实验研究和发现表明,反应.扩散模型的许多假设是过于理想化的,模型的更为合理化是将来研究的重点【4】。
2.2 Capdeville 生物膜增长动力学模型【4,5】
20世纪90年代初,法国CapdeviUe 教授所领导的实验室提出生物膜反应器活性物质和非
活性物质的概念,并在此基础上建立了新的生物膜增长动力学模型。
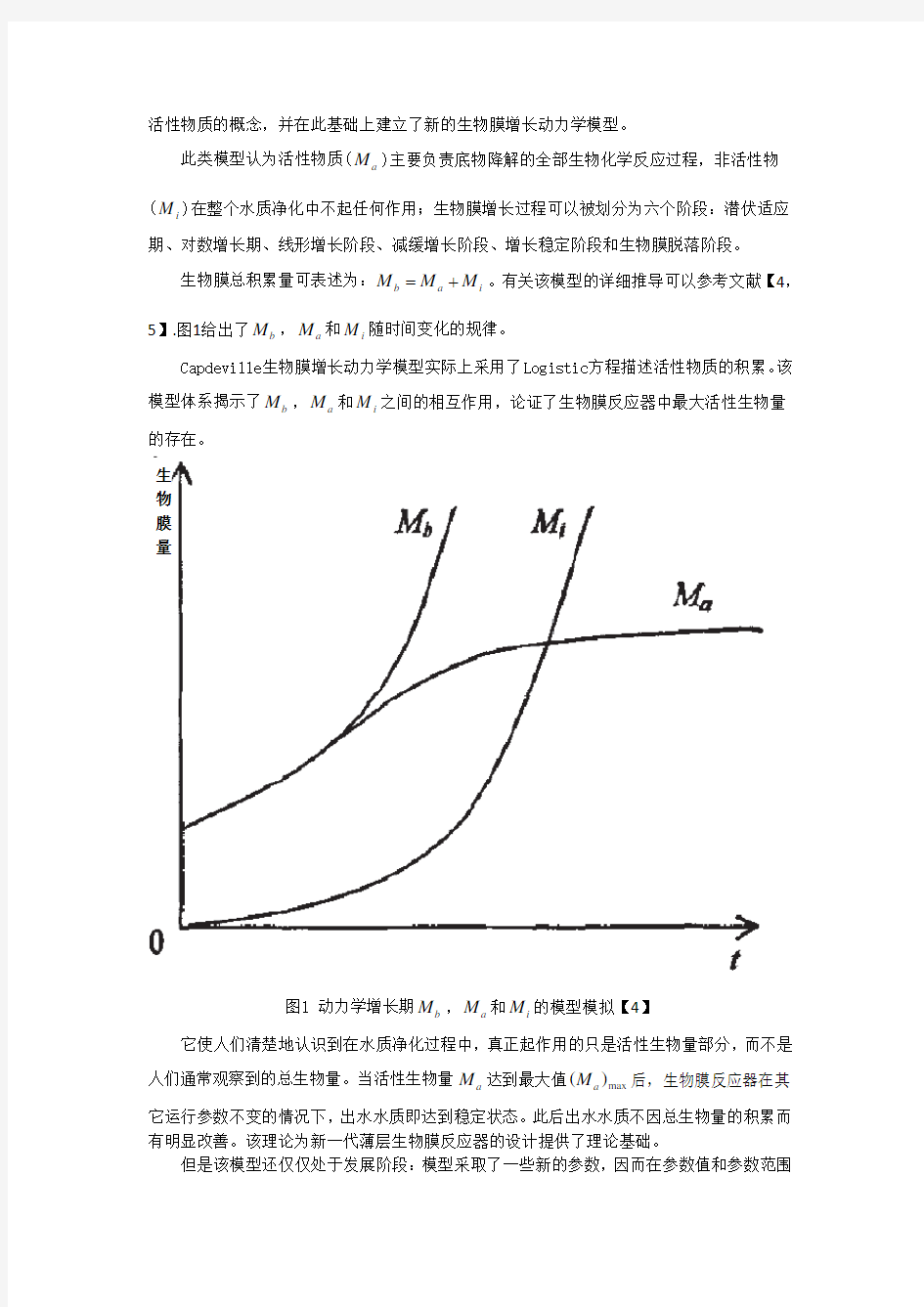
此类模型认为活性物质(a M )主要负责底物降解的全部生物化学反应过程,非活性物(i M )在整个水质净化中不起任何作用;生物膜增长过程可以被划分为六个阶段:潜伏适应期、对数增长期、线形增长阶段、减缓增长阶段、增长稳定阶段和生物膜脱落阶段。 生物膜总积累量可表述为:b a i M M M =+。有关该模型的详细推导可以参考文献【4,5】.图1给出了b M ,a M 和i M 随时间变化的规律。
Capdeville 生物膜增长动力学模型实际上采用了Logistic 方程描述活性物质的积累。该模型体系揭示了b M ,a M 和i M 之间的相互作用,论证了生物膜反应器中最大活性生物量的存在。
图1 动力学增长期b M ,a M 和i M 的模型模拟【4】
它使人们清楚地认识到在水质净化过程中,真正起作用的只是活性生物量部分,而不是人们通常观察到的总生物量。当活性生物量a M 达到最大值max ()a M 后,生物膜反应器在其它运行参数不变的情况下,出水水质即达到稳定状态。此后出水水质不因总生物量的积累而有明显改善。该理论为新一代薄层生物膜反应器的设计提供了理论基础。
但是该模型还仅仅处于发展阶段:模型采取了一些新的参数,因而在参数值和参数范围生
物
膜
量
的确定上,还没有很成熟的工作成果可以采用;在实验中活性物质和非活性物质很难区分,因此尽管间接验证是成立的,但直接验证还相当困难;此外,模型没有考虑脱落、外部环境等系统条件对生物膜形成和动力学特征的影响。这一切均表明该模型还有很多需要将来去完善的地方。
2.3 元胞自动机(CA)模型
许多学者基于元胞自动机(cellular automaton)的概念,结合描述生物膜内传质和反应的微分方程,进行数值模拟,研究系统参数对生物膜结构、形态和功能的影响;或将生物膜细胞生长、衰亡、脱落、基质扩散和基质利用考虑为随机过程,假定简单的局部的规则,建立生物膜形成和结构的2D和3D数学模型,研究各个参数对生物膜结构和形态的影响。
Colasanti【6】在1992年首先将元胞自动机概念作为一种数学方法引入生物膜建模中以描述生物膜的异质性(heterogeneity)。Wimpenny【7】在回顾了有关微生物生物膜基本结构的文献后发现至少存在三种概念性的模型:异质镶嵌型生物膜模型、水通道生物膜模型和紧密生物膜模型。他认为在考虑生长资源的影响后,三种模型可能都是正确的,其差异只是由于生物膜内的不同基质浓度造成的;他通过细菌定殖的实验研究和利用元胞自动机所作的数值模拟证实:生物膜结构主要取决于基质浓度。
Hermanowiez【8-10】通过数值模拟发现,浓度边界层和水力边界层的厚度对生物膜的结构有重要的影响,液相主体营养物浓度的影响并不重要。当外部传质限制明显时,模型生物膜发展为一个开放结构;当浓度边界层变薄且外部传质加强时,产生一层致密的生物膜。Hermanowiez为简单的局部规则控制细胞的生长和脱落,而细胞的自组织产生了生物膜结构的不同形态【9,10】。
也有学者利用元胞自动机模型研究生物膜3D异质性及种群动力学。例如:Picioreanu 等人【11】研究水力条件对生物膜的松动、脱落的影响;Rittmann等人【12】主要研究生物膜边界层的厚度和形状对生物膜的结构及活性的影响;Noguera等人【13】主要描述厌氧生物膜的异质性和微生物种群的形成对生物膜3D结构的影响。
Laspidou和Rittmann【14,15】引入了生物膜压实效应(consolidation),建立了一个包括活性物质、非活性物质和胞外聚合物(EPS)等在内的统一的多组分元胞自动机模型(UMCCA),并通过数值模拟研究生物膜密度、各组分的变化以及生物膜形态,并和已有的实验进行了比较。
以上提到的有关生物膜的元胞自动机模型,基本上都可以看作是一种混合模型,他们将描述物理和生化过程的微分方程和元胞自动机模型结合起来,主要利用元胞自动机算法对微分方程进行求解。Pizarro等人【16】采用了新的方法,建立了一个的完全意义上的CA模型,将生物膜生长中出现的许多现象,如基质扩散、基质利用、生物膜脱落和衰亡等都利用随机过程(元胞自动机)来模拟,并利用此模型来预测基质梯度和通量,得到了与微分方程模型相似的结果,且在高浓度时更加精确。
元胞自动机模型在生物膜动力学建模以及在生物膜形态和结构模拟方面得到了广泛应用,并取得了许多有意义的成果。不过由于元胞自动机是通过人为假定的计算机算法规则来反映动力学,本身并没有很清楚的物理背景,因此元胞自动机模型的理论基础及其有效性受到很大的质疑。元胞自动机模型应在未来的研究中不断完善。
2.4 多物种复合生物膜模型
生物膜内除存在异养菌以外通常同时存在硝化菌和反硝化菌等自养菌。因此多物种模型是生物膜建模过程的研究热点。但由于以下几点原因,使得多物种生物膜的建模和模拟成为废水处理的生化操作建模中最为复杂和最为困难的过程之一。首先,必须同时考虑反应和传质。第二,不同物种会同时竞争一种营养物质。第三,不同物种会在生物膜内部竞争生长空间。生物膜内多物种建模的最理想方式是假定生物膜内任一点处都可以生长各种类型的细
菌,而它们的最终分布决定于竞争到的营养和生长空间。限于篇幅的原因,我们在这里对多物种生物膜建模仅作简单介绍。具体可以参考《Biofilms》中的第1l章【17】,或其它关于此类建模的论文【18-26】。此外,Reichert建立了一个应用计算机语言处理一般用途的多物种生物膜模型的软件AQUASIMl【27】(或参考http://www.aquasim.eawag.ch)。
Wanner和Gujer【19】研究了异养菌和自养菌对液相主体内溶解氧的竞争问题,并考察了共存条件。Rittmann和Manem【22】建立了仅仅竞争生长空间的多种细菌的稳态生物膜模型。物种的最终分布将决定于其在生物膜内各点处的相对生长速度。Rittmann等【23】在2002年提出了瞬态多物种生物膜模型(TSMSBM),该模型综合了已有模型的重要特性,用以描述时变条件下特别是反冲洗引起的周期性脱落情况下的多物种生物膜。
Cao等【25】发展了一个外电流作用下的多物种生物膜内稳态模型,用于描述异养菌和自养菌的相互竞争。王志盈等[26]根据活性生物膜扩张原理,建立了硝化生物膜中微生物相互作用的非稳态解析数学模型,并可预测生物膜厚度的变化与微生物菌群的空间分布。
综上所述,反应,扩散模型主要关心基质的去除动力学,Capdeville生物膜增长动力学模型主要关心活性生物量和非活性生物量的积累,而原胞自动机模型则主要关心生物膜的形态和结构,复合生物膜模型则更关心生物膜内各微生物种群的相互作用和空间分布。这四类模型形成了生物膜动力学理论研究的主流。
3.生物膜动力学的实验研究
除了理论模型上的进展外,最近十几年科学家们开展了不少针对生物膜的实验研究,做出了许多发现,其中最重要的发现是生物膜的异质性(heterogeneity)和分形结构(fraetal structure)。这些新的实验发现已经明显地改变了人们对生物膜系统的看法,并引起了对生物膜动力学理论模型的重要修正。
3.1 生物膜的异质性
Wanner【28】在总结了许多新的实验发现指出:溶解性物质在生物膜内的传质并不总是通过分子扩散;不能排除与生物膜内物种净增长速率相关的颗粒物质的传递;生物膜内的液相体积分数(孔隙率)不是常数;生物膜表面的细胞和颗粒的脱落和吸附是重要的过程。Wimpermy等【29】在2000年总结了最近在实验方面关于生物膜形成、结构方面的新发现,也指出了生物膜结构的异质性。
许多文献【29-31】已经指出,受多种因素的影响,生物膜结构并非是各向同性的。Gialtema【30】指出,生物膜的结构受微生物类型、生长表面、液相流动类型、反应器类型和生物膜脱落等因素的影响。Beer等【31】和Massk-Deya等【32】都观察到多种群共生的生物膜(好氧膜和厌氧膜)中存在着孔(cavity)、空隙(void)及通道(channel)。因此,在建立生物膜数学模型时应该考虑三维方向上的传质,而不仅仅是考虑垂直于载体表面方向的传质。
Beer等【33】和Stoodly等【34】认为生物膜是由不同的通道隔离开的质量分布均匀的多孔结构,类似一个海绵体。Tijhuis等【35】认为生物膜的形成是一个动态过程,生物膜在外力作用下迅速并连续不断地生成裂缝,这些裂缝又不断地被新生长出的微生物填满。3.2 生物膜结构参数的测定
由于上述实验事实的发现,传统生物膜的结构参数如生物膜厚度、密度、孔隙率等已不能准确地描述生物膜的形态和结构。而“通道(channel)”、“孔(cavity)”、“粗糙的(rough)”等词语只能用来定性地描述生物膜形态和结构【31,32】。因此需要有新的生物膜结构参数和相应的定量化方法来定量地描述生物膜的形态。
最近十几年以来,显微实验技术特别是激光扫描共聚焦显微镜(Confocal Laser Scanning Microscope,CLSM)技术得到了飞速发展,并在生物膜的研究中得到了广泛应用。王荣昌等【36】介绍了激光扫描共聚焦显微镜(CLSM)成像系统的构成和工作原理,并对其应
用研究生物膜的微生物群落结构和空问构型的一些实例进行了综述,同时指出了其应用于生物膜研究的优、缺点及今后的发展前景。激光扫描共聚焦显微镜(CLSM)也经常同荧光原位杂交技术(FISH)联合使用,用于复合生物膜内种群变化和空问分布的实验观测【37-39】。
激光扫描共聚焦显微镜(CLSM)和计算机图像分析技术的发展,使得研究者能够将生物膜结构可视化,在此基础上建立新的生物膜的结构参数及其实验测定方法,作为传统生物膜结构参数的补充【40,42,45】。
Lewandowski等【40】和Jackson等【41】通过实验定量地测定了生物膜的四个结构参数:组织熵(textural entropy),面孔隙度(areal porosity),分形维(fractal dimension),最大扩散距离(maximum diffusion distance)。文献【42】具体地将该方法从二维图像处理扩展到了三维,并给出了从三维激光扫描共聚焦显微镜图像计算生物膜分形维数的具体算法。同时文献【42】也建立了其它许多定量描述生物膜三维结构参数的测量方法,而且讨论了每一个参数的具体含义,如平均扩散距离(average diffusion distance,ADD)和最大扩散距离(maximum diffusion distance,MDD),在x、Y、z三个方向上的平均运动长度(average run lengths)、纵横比(aspect ratio)、能量和异质性。
曹宏斌等【43】研究发现,当没有外电流作用时,自养、异养/自养生物膜的孔隙率从表层向内层逐步减少,分别由表层的大约92%和96%逐步减少到最内层的54%左右;而分形维分别由表层的1.1和1.05逐步增大,到最内层时达到1.4;随着通过反应器的电流密度增大,自养、异养/自养生物膜表层的空隙率将略有减少,而分形维数略有增大。
Hermanowiez等人【44,45】测定了异养好氧生物膜分形维。此分形维是通过计算像素与像素关联函数的斜率得到的。他们通过这种方法确定了两个尺度上的分形维。在小尺度上(小于3um)生物膜的分形维为2.94,接近于拓扑维。在较大尺度上(12~30um)分形维是相当小的(2.50~2.67)。在较大的尺度上,平行于水流方向测得的分形维比垂直于水流方向测得的分形维大得多。
此外,Moghabghab[46]定义并实际测定了生物膜的两个分形维数:描述表面粗糙度的边界分形维数(boundary fraetal dimension),其值在1.02~1.34之间;描述孔隙率的Sierpinski分形维(Sierpinski fraetal dimension),其值在1.932~1.974之间。这两个分形维不受取样时间、生物膜厚度和孔隙率以及空间位置的影响。
从现有的实验结果来看:生物膜的分形维数的数值相差较大,一方面可能与实验测定的方法、取样的部位(生物膜的深度)、以及生物膜生长的时间和生物膜的性质等因素有关;另一方面,则有可能是由于采用了具有不同定义和含义的分形维数造成的。在不同的尺度大小和不同的方向上,分别对应着不同大小的分形维数,一方面进一步说明了生物膜的异质性,另一方面也说明了生物膜内存在着多个尺度和多个层次,因此要完整的描述生物膜的结构和形态,需要引入新的结构参数,如多重分形等。不同研究人员测定分形维数及其他一些形态和结构参数的方法有差异,因此,要用定量参数完整地描述生物膜的形态和结构,并使这些参数值具有可比性,必须建立统一的标准化的测定方法,也可能需要引入新的结构参数,如多重分形等。
4 结语——生物膜动力学研究的展望
生物膜体系是一个远离平衡态的、开放的热力学系统,涉及多种组分、多个物种的复杂的动力学过程,形成了复杂、有序的网络结构。因此要想合理地研究和描述生物膜形成、生长和稳定过程,必须从非线性物理着手,充分应用分形理论、自组织理论、耗散结构等复杂系统理论,从全新的角度和视点来阐明生物膜的形成、发展、结构和功能。
其次,从生物数学的角度看,生物膜本质上是多基质、多种群的包含竞争、捕食关系的一种多层次的生物网络结构【2-4】。然而迄今为止,很少有人从生物数学的角度去研究生物膜内各微生物种群之间的竞争和捕食关系,及其对微生物种群在载体上的分布和生物膜的
生长、形成、结构和功能的影响。因而,必须从微观生态入手,加强这方面的研究。
最后,我们必须充分认识到,生物膜动力学数学建模、数值模拟与实验研究是紧密联系、密不可分的。数学模型提供的是评估假设前提合理性的方法与确定合理的参数估计方法。实验观察揭示的是模型假设的合理性,以及提供一些用于模拟和反应器设计的参数值和参数范围。只有数学模型与实验研究紧密地结合起来,我们才能更加深刻地理解生物膜的形成、形态、结构和功能,也才能将其用于指导我们的实践,提高生物膜反应器的处理效率。
参考文献
【1】Noguera DR,Okabe S,Picioreanu C,Biofilm modeling:present status and future directions,Water Science and Technology,1999,39(7):273~278.
【2】 Grady Jr CPL,Daigger GT,Lim HC,Biological Wastewater Treatment:Revised and Expanded.2nd ed.New York:Marcal Dekker,Inc,1999.72l~747,812,763-840.
【3】格雷迪等著,张锡辉,刘勇弟译,废水生物处理:改编和扩充.第二版.北京:化学工业出版社.2002.471~484.
【4】刘雨,赵庆良,郑兴灿编著.生物膜法污水处理技术.北京:中国建筑工业出版社,2000.56~73.
【5】刘雨,王岐东,生物膜增长动力学模型,北京轻工业学院学报,1997,15(2):28~32.
【6】 Colasanti R,Cellular automata models of microbial colonies,Binary,1992 4:191-193.
【7】 Wimpenny JWT,Colasanti&A unifying hypothesis for the structure of microbial biofilms based on cellular automaton models,FEMS Microbiology Ecology,1997,22(1):1~16.
【8】 Hermanowicz SW,A model of two-dimensional biofilm morphology,Water Science and Technology,1998,37(4—5):219~222.
【9】 Hermanowicz SW.Two-Dimensional simulation ofbiofilm development:effects ofexterllal environmental conditions,Water Science and Technology,1999,39(7):107~114.
【10】 Hcrmanowicz SW,A simple 2D biofilm model yields a variety ofmorphological features,Mathematical Biosciences,2001,169(1):l~14.
【11】 Picioreanu C,van Loosdreeht MC.M,Heijnen JJ,Discrete-differential modeling ofbiofilm slrueture,W aterScience and Technology,1999,39(7):115~122.【12】 Rittmann BE,Pettis M,Reeves HW,et a1.,How biofilm clusters affect substrate flux and ecological selection,Water Science and Technology,1999.39(7):
99-105.
【13】 Noguera DR, Pizarro G Stahl D A,et a1.,Simulation of multi—species biofilm development in three dimensions,Water Science and Technology,1999,39(7):123~130.
【14】 Laspidou CS,Rittmann BE,Modeling the development of biofilm density including active bacteria,inert biomass,and extracellular polymeric
substances,Water Research,2004,38(14一15):3349.3361.
【15】 Laspidou CS,Rittmatm BE,Evaluating trends in biofilm density using the UMCCA model,WaterResearch,2004,38(14—15):3362~3372.
【16】 Pizarro G,Griffeath D,Noguera DR Quantitative cellular automatic for
biofilms.Journal of Environment engineering,2001.127(9):782—789.【17】 Gujer W,Wanner O,Modeling mixed population biofilms.In:Characklis W G and Marshall K C,eds.Biofilms.NewYork:Wiley,1990.397—443.
【18】 Kissel JC,McCarty PL&Street RL,Numerical simulation of mixed-culture biofilm,Journal ofEnvironmental Engineering,1984,110(2):393~411.【19】 Wanner O,Gujer Wj Competition in biofilms,Water Science and Technology, 1984,17(2-3):27,44.
【20】 Wanner O,Gujer W:A multispecies biofilm model,Biotechnology and Bioengineering,1986,28(3):314~328.
【21】 Wanner O,Reichert P.Mathematical modeling of mixed-culture biofilms,Biotechnology and Bioengineering,1996,49(2):172~184.
【22】 Rittmann BE,Manem JA. Development and experimental evaluation of a steady-state,multispecies biofilm model. Biotechnology and
bioengineering,1992,39(9):914~922.
【23】 Rittmann BE,Stilwell D,Ohashi A,The transient-state,multiple—species biofilm model for biofiltration processes,Water Research,2002,36(9):
2342~2356.
【24】 Rauch W;Vanhooren H,Vanrolleghem PA,A simplified mixed-culture biofilm model,Water Research,1999,33(9):2148-2162.
【25】 Cao HB,Li XG,Wu JC,et a1.,Simulation ofthe effects ofdirect electric current on multispecies biofilms,Process Biochemistry,2003,38(8):l139~1145.
【26】王志盈,彭党聪,袁林江等,低溶解氧下硝化生物膜中菌群的竞争增殖模型。
中国给水排水,1999.15(12):18-22.
【27】 Reichert P.AQUASIM,a tool for simulation and data analysis in water and wastewatertreatment,WaterScience and Technology,1994,30(4):21-30.【28】 Wanner O,New experimental findings and biofilm modelling concepts,Water Science and Technology,1995,32(8):133-140.
【29】 Wimpenny J,Manz W,Szewzyk U,Heterogeneity in biofilms。FEMS Microbiology Reviews,2000,24:661—671.
【30】 Gialtema A'Arts PAM,vail Loosdrecht MCM,Heterogeneity of biofilms in rotating annular reactors:OCCUlTeUCe,structure and consequences,
Bioteehnology and Bioengineering,1 994,44(2):1 94--204.
【31】 De Beer D,Stoodley B Roe F'et a1.,Effects of biofilm structures on oxygen distribution and mass transfer,Biotechnology and Bioengineering,1994,43(1
1):1131-1138.
【32】 Massol·Deya AA,Whallon J,Hickey RF,et a1.,Channel structures in aerobic biofilms of fixed-film reactors treating contaminated groundwater,Applied Environmental Microbiology,1995,61(3):769~777.
【33】 De Beer D,Stoodley P,Lewandowski Z,Liquid flow in heterogeneous biofilms,Biotechnology and Bioengineering,1994,44(5):636~641.
【34】 Stoodley P,De Beer D,Lewandowski Z,Liquid flow in biofilm system,Applied EnvironmentalMicrobiology,1994,60f8):2711-2716.
【35】 Tijhuis L,Van Bentham WAJ,Van Loosdrecht MCM,ct a1.。Solids retention
time in spherical biofilms in a biofilm airlift suspension
reactor,Biotechnology and Bioengineering,1994,44(8):867~879.
【36】王荣昌,文湘华,钱易,激光扫描共聚焦显微镜用于生物膜研究,中国给水排水,2003,19(12):23,25.
【37】 Falkentoft CM,Muller E,Amz P,et a1.,Populmion changes in a biofilm reactor for phosphorus removal as evidenced by the use of FISH,Water Research,
2002,36(2):491~500.
【38】 Persson F,Wik T,Sfrensson F,et a1.Distribution and activity of ammonia oxidizing bacteria in a large full-scale trickling filter,Water Research,2002,36(6):1439-1448.
【39】 Thurnheer T,Gmfiir Guggenheim B,Multiplex FISH analysis ofa six·species bacterial biofilm,Journal of Microbiological Methods,2004,56(1):37—7.【40】 Lewandowski Z,Webb D,Hamilton M,et a1.,Quantilying biofilm structure,Water Science and Technology,1999,39(7):71~76.
【41】 Jackson G,Beyenal H,Rees WM,et a1.,Growing reproducible biofilms with respect to structure and viable cell counts,Journal ofMicrobiological
Methods,2001,47(1):I-10.
【42】 Beyenal H,Donovan C,Lewandowski Z,et a1.,Three-dimensional biofilm structure quantification,Journal of Microbiological Methods,2004,59(3):395--413.
【43】曹宏斌,姜斌,李鑫钢等,外电流对生物膜空隙率和分形维数分布的影响,环境科学,2003,24(4):85-88
【44】 Hermanowicz SW,Schindlel"U.Wilderer P.Fractal slxucture of biofilms:new tools for investigation of morphology,WaterScience and Technology,1995,32(8):
99-105.
【45】 Hermanowicz SW,Schindler U,Wilderer P’Anisotropic morphology and fractal dimemions ofbiofilms,Water Research,1996,30(3):753~755.
【46】 Moghabghab Ran]a,External surface and porosity of RBC biofilms in leaehate pre-treatment:[Master].Toronto:University ofToronto,1997.1~64 【47】陈黎明,柴立和. 生物膜废水处理系统的数学模型及机理探讨. 自然科学进展,2005,15(7):843~848
生物膜的研究进展
第7卷第5期1998年10月 环境科学进展 ADVANCESINENVIRONMENTALSCIENCE Vol.7,No.5 Oct.,1999生物膜的研究进展Ξ 王文军1、2 王文华1 黄亚冰1 张学林2 (1中国科学院生态环境研究中心环境水化学国家重点实验室,北京100085) (2中国科学院长春地理研究所,长春130021) 摘 要 本文综述了近年来生物膜研究成果,包括生物膜的发育形成、形态结构、组成、物理-化学特征、抗性等;生物膜在污水处理方面的作用和微生物组织腐蚀性的负效应。 关键词:生物膜 特征 作用 生物膜在天然水环境中和工程处理过程中起着重要的作用[1-3]。在天然水环境中,绝大部分矿物颗粒表面覆盖着有机外壳[4],这些有机外壳由腐殖酸物质和生物膜组成,它们将强烈地改变矿物颗粒的吸附行为,这种表面吸附作用在河水污染物的迁移过程中起着决定性作用。在工业应用中,生物膜的作用表现在废水处理,以及酸性矿物排泄物的生物修复等方面,例如在水和废水处理系统中,生物膜反应器比悬浮生长反应器具有更大的优势,能提高生物量在反应器中的滞留程度,促进对污染物降解效率。生物膜的破坏性作用表现在清洁水系统中,以及微生物诱导的腐蚀等方面[5,6]。随着对生物膜在自然环境(如水、土、生物环境)中和工业应用方面的重要性的不断认识,在过去的二十多年,人们对生物膜的兴趣极大地增加[7]。美国、德国、日本、英国、法国等国家对生物膜进行了大量的研究[1-31],取得了一些初步的研究成果。 一、生物膜的形成及影响因素 生物膜形成于自然环境和人工环境中。在自然环境条件下,生物膜存在于几乎所有暴露于水中的固体表面上,代表了一类微生物群体,其中有各种寄居者如固着细菌、原生动物、真菌和藻类[4-9]。这些微生物细胞及非生物物质镶嵌在微生物分泌的有机聚合物基质(Matrix)中,聚合物基质由细菌胞外聚合物质和腐殖质等其它有机物质组成。即生物膜代表了一种稳定的由微生物细胞组成的复杂混合物的微生态系统,细胞镶嵌在胞外聚合物的基质中,并且附着到固体表面。生物膜发育形成的条件和时间序列大致为[9]: (1)存在着清洁的可用于聚居的固体表面;(2)一种有机分子膜快速形成;(3)聚结的细胞 Ξ1国家自然科学基金资助项目:29777027 2中国科学院武汉水生所淡水生态与生物技术国家重点实验室开放基金资助
细菌生物膜研究进展 (1)
306 中国医学文摘耳鼻咽喉科学 NEWS AND REVIEWS/November 2009, Vol.24, No.6 专题论坛 抗生素的合理应用 EATURE 1 生物膜的概念 细菌生物膜是指在多聚糖、蛋白质和核酸等组成的基质内相互粘连粘附于物体表面的细菌群体[1]。生物膜可以由一种或几种细菌混合生长而成。乳酸乳球菌与萤光假单胞菌混合形成的生物膜就是一个典型的例子。乳酸乳球菌自身不易形成生物膜,但可以提供给萤光假单胞菌乳酸作为养料,而萤光假单胞菌帮助乳酸乳球菌固定在物体表面,并且消耗氧气为乳酸乳球菌这一厌氧菌提供更合适的生长环境[2]。 生物膜的生命周期分为附着、生长和分离3部分。附着阶段,物体表面的血清蛋白和其他物质作为连接物介导细菌的附着;生长阶段,细菌通过分裂并在物体表面定植,生成聚合物基质,使得生物膜形成三维结构,并形成隧道,这些隧道帮助营养物质的交换以及废物的排出,并调节生物膜内的pH 值。生物膜中的细菌对氧气和营养的需要有所减少,废物通过其内的管道得以排出。生物膜内细菌间的紧密接触为携带耐药基因的质粒的交换和对密度感应分子的交流提供了良好环境。生物膜内的细菌间更利于质粒、酶和其他分子的交换,通过化学信号进行交流。生物膜的形成需要细菌间的化学信号进行协调。使得细菌能感知到周围细菌的存在并对环境变化作出相应的反应。这一过程称为密度感应(quorum-sensing )。虽然不同细菌的生物膜有其特异性,但均具有一些普遍的结构特征。生物膜中细菌形成的微菌落间具有间隙空位(interstitial voids ),液体可在这些间隙中流动,使得营养物质、气体和抗菌药物得以扩散。生物膜的结构随着外部和内部的改变而持续变化。 2 生物膜与临床 99%的细菌以生物膜的形式生活,美国疾病控制与预防中心估计至少65%的人类细菌感染与生物膜有关[3]。生物膜已经被证实与慢性中耳炎、中耳胆脂瘤、慢性腺样体炎[1]等疾病相关。Pawlowski 等[4]于2005年在耳蜗植入体上发现了细菌生物膜。Cryer 等[5]于2004年发现一些慢性鼻窦炎手术治疗后症状仍持续 细菌生物膜研究进展 郑波 [关键词] 生物膜(Bio ?lms );抗药性,细菌(Drug Resistance ,Bacterial ) 郑波 北京大学第一医院临床药理研究所,北京 100034 广东人,副教授,副主任医师,主要从事细菌耐药机制和抗菌药物合理应用的研究工作。Email :doctorzhengbo@https://www.sodocs.net/doc/3619150965.html, 的患者鼻窦中存在生物膜,这些患者主要为铜绿假单胞菌感染。Ramadan 等[6]于2005年对5位慢性鼻窦炎患者进行黏膜活检,对标本进行扫面电镜检查均发现有生物膜的存在。此外,生物膜已被证实与下列感染有关:慢性前列腺炎、导管相关感染、人工关节感染、牙周病、心内膜炎以及囊性纤维化患者的假单胞菌肺炎等。 3 生物膜与抗菌药物耐药 生物膜内细菌对抗菌药物的敏感性较游离状态时显著降低,最低可降低1000倍。其原因包括生物膜的结构阻止了药物的传输或生物膜中的细菌的生理学改变等。以前一直认为生物膜介导的对抗菌药物耐药的原因是抗菌药物难以渗透入生物膜。但一些研究否认了这一假设。研究显示喹诺酮类可以很快的渗透到铜绿假单胞菌和肺炎克雷白杆菌生物膜的深部[7,8],四环素可很快的渗透到大肠埃希菌生物膜内,万古霉素可以很快渗透到表皮葡萄球菌生物膜内。目前唯一得到证实的是氨基糖苷类药物,由于生物膜中的基质带负电荷,而氨基糖苷类带有正电荷,因此氨基糖苷类药物难以渗透到生物膜的深部[9]。 生物膜对β内酰胺类耐药性增加的机制之一是细菌产生的β内酰胺酶在生物膜表面基质内聚集,可达到很高的浓度,能迅速的将渗透进生物膜内的β内酰胺类抗生素水解掉,有效保护深部细菌不被β-内酰胺类抗菌药物灭活[10]。有研究证实氨苄西林会被肺炎克雷白杆菌生物膜表层中聚集的β内酰胺酶快速水解。 生物膜造成的缺氧环境也增加了对抗菌药物耐药性。一项在囊性纤维化患者生成的铜绿假单胞菌生物膜的研究显示氧气仅能渗透到生物膜的25%深度。铜绿假单胞菌在厌氧条件下比在有氧条件下对抗菌药物的敏感性明显降低[11]。 由于很多抗菌药物对繁殖期细菌杀伤作用更强大,如青霉素类、头孢菌素类和碳氢霉烯类等。在生物膜深部的细菌受氧气、营养物质缺乏的影响及可能存在的密度感应系统的调控,使得细菌的生长、繁殖速度下降,影响抗菌药物对其作用。因此在抗菌药物作用下,生物膜中相对敏感的细菌会被杀死,但耐药菌会持
生物信息学的主要研究内容
常用数据库 在DNA序列方面有GenBank、EMBL和等 在蛋白质一级结构方面有SWISS-PROT、PIR和MIPS等 在蛋白质和其它生物大分子的结构方面有PDB等 在蛋白质结构分类方面有SCOP和CATH等 生物信息学的主要研究内容 1、序列比对(Alignment) 基本问题是比较两个或两个以上符号序列的相似性或不相似性。序列比对是生物信息学的基础,非常重要。两个序列的比对有较成熟的动态规划算法,以及在此基础上编写的比对软件包BLAST和FASTA,可以免费下载使用。这些软件在数据库查询和搜索中有重要的应用。 2、结构比对 基本问题是比较两个或两个以上蛋白质分子空间结构的相似性或不相似性。已有一些算法。 3、蛋白质结构预测,包括2级和3级结构预测,是最重要的课题之一 从方法上来看有演绎法和归纳法两种途径。前者主要是从一些基本原理或假设出发来预测和研究蛋白质的结构和折叠过程。分子力学和分子动力学属这一范畴。后者主要是从观察和总结已知结构的蛋白质结构规律出发来预测未知蛋白质的结构。同源模建(Homology)和指认(Threading)方法属于这一范畴。虽然经过30余年的努力,蛋白结构预测研究现状远远不能满足实际需要。 4、计算机辅助基因识别(仅指蛋白质编码基因)。最重要的课题之一 基本问题是给定基因组序列后,正确识别基因的范围和在基因组序列中的精确位置.这是最重要的课题之一,而且越来越重要。经过20余年的努力,提出了数十种算法,有十种左右重要的算法和相应软件上网提供免费服务。原核生物计算机辅助基因识别相对容易些,结果好一些。从具有较多内含子的真核生物基因组序列中正确识别出起始密码子、剪切位点和终止密码子,是个相当困难的问题,研究现状不能令人满意,仍有大量的工作要做。 5、非编码区分析和DNA语言研究,是最重要的课题之一 在人类基因组中,编码部分进展总序列的3~5%,其它通常称为“垃圾”DNA,其实一点也不是垃圾,只是我们暂时还不知道其重要的功能。分析非编码区DNA 序列需要大胆的想象和崭新的研究思路和方法。DNA序列作为一种遗传语言,不仅体现在编码序列之中,而且隐含在非编码序列之中。 6、分子进化和比较基因组学,是最重要的课题之一 早期的工作主要是利用不同物种中同一种基因序列的异同来研究生物的进化,构建进化树。既可以用DNA序列也可以用其编码的氨基酸序列来做,甚至于可通过相关蛋白质的结构比对来研究分子进化。以上研究已经积累了大量的工作。近年来由于较多模式生物基因组测序任务的完成,为从整个基因组的角度来研究分子进化提供了条件。 7、序列重叠群(Contigs)装配 一般来说,根据现行的测序技术,每次反应只能测出500或更多一些碱基对的序列,这就有一个把大量的较短的序列全体构成了重叠群(Contigs)。逐步把它们拼接起来形成序列更长的重叠群,直至得到完整序列的过程称为重叠群装配。拼接EST数据以发现全长新基因也有类似的问题。已经证明,这是一个NP-完备
生物信息学现状与展望
研究生课程考试卷 学号、姓名: j20112001 苗天锦 年级、专业:2011生物化学与分子生物学 培养层次:硕士 课程名称:生物信息学 授课学时学分: 32学时 2学分 考试成绩: 授课或主讲教师签字:
生物信息学现状与展望 摘要:生物信息学是一门新兴学科,起步于20世纪90年代,至今已进入"后基因组时代",本文对生物信息学的产生背景及其研究现状等方面进行了综述,并展望生物信息学的发展前景。生物信息学的发展在国内、外基本上都处在起步阶段。 关键词:生物信息学;生物信息学背景;发展前景 一、生物信息学概述 1.生物信息学发展历史 随着生物科学技术的迅猛发展,生物信息数据资源的增长呈现爆炸之势,同时计算机运算能力的提高和国际互联网络的发展使得对大规模数据的贮存、处理和传输成为可能,为了快捷方便地对已知生物学信息进行科学的组织、有效的管理和进一步分析利用,一门由生命科学和信息科学等多学科相结合特别是由分子生物学与计算机信息处理技术紧密结合而形成的交叉学科——生物信息学(Bioinformatics)应运而生,并大大推动了相关研究的开展, 被誉为“解读生命天书的慧眼”【1】。 研究生物细胞的生物大分子的结构与功能很早就已经开始,1866年孟德尔从实验上提出了假设:基因是以生物成分存在。1944年Chargaff发现了著名的Chargaff规律,即DNA中鸟嘌呤的量与胞嘧定的量总是相等,腺嘌呤与胸腺嘧啶的量相等。与此同时,Wilkins与Franklin用X射线衍射技术测定了DNA纤维的结构。1953年James Watson 和FrancisCrick在Nature杂志上推测出DNA 的三维结构(双螺旋)。Kornberg于1956年从大肠杆菌(E.coli)中分离出DNA 聚合酶I(DNA polymerase I),能使4种dNTP连接成DNA。Meselson与Stahl (1958)用实验方法证明了DNA复制是一种半保留复制。Crick于1954年提出了遗传信息传递的规律,DNA是合成RNA的模板,RNA又是合成蛋白质的模板,称之为中心法则(Central dogma),这一中心法则对以后分子生物学和生物信息学的发展都起到了极其重要的指导作用。经过Nirenberg和Matthai(1963)的努力研究,编码20氨基酸的遗传密码得到了破译。限制性内切酶的发现和重组DNA的克隆(clone)奠定了基因工程的技术基础【2】。自1990年美国启动人类基因组计划以来,人与模式生物基因组的测序工作进展极为迅速。迄今已完成了约40多种生物的全基因组测序工作,人基因组约3x109碱基对的测序工作也接近完成。至2000年6月26日,被誉为生命“阿波罗计划”的人类基因组计划终于完成了工作草图,预示着完成人类基因组计划已经指日可待。生物信息学已成为整个生命科学发展的重要组成部分,成为生命科学研究的前沿。 2.生物信息学研究方向 2.1 序列比对
支原体生物膜研究进展_叶晓敏
·综述·支原体生物膜研究进展 叶晓敏,陆春 (中山大学附属第三医院皮肤科,广东广州510630) [摘要]近几年,支原体生物膜研究逐渐受到研究人员的关注。多种支原体都被证实具有生物膜形成 能力,生物膜形成后支原体耐药性增加,研究生物膜对于防治临床支原体感染有着重大意义。本文从 目前报道的几种支原体生物膜的形成及结构、生物膜形成的影响因素、生物膜形成对支原体药物敏感 性的影响及可能机制等几个方面综述了目前对支原体生物膜的研究进展。 [关键词]支原体;生物膜 [中图分类号]R759[文献标识码]A[文章编号]1674-8468(2011)01-0060-04 生物膜(Biofilm,BF)是微生物在生长过程中附着于物体表面而形成的由微生物的细胞及其分泌的聚合物等所组成的膜样多细胞复合体[1]。生物膜的存在可以增强病原微生物对宿主免疫攻击及抗菌药物的抵抗力。目前对大量支原体的研究已发现很多支原体都具有形成生物膜的能力。生物膜形成后增强了支原体对环境压力如热、干燥、缺氧、高渗透压等[2-3]及对抗菌药物的抵抗力[4]。本文从支原体生物膜的形成及结构、生物膜形成的影响因素、生物膜形成对支原体药物敏感性的影响及可能机制等几个方面对目前支原体生物膜的研究进展作一综述。 1支原体生物膜的鉴定及其形成和结构 生物膜是微生物细胞不断粘附、聚集,并包裹在自身生成的胞外基质中形成的多聚复合物,体积上15%由细胞组成,85%由胞外基质组成。目前生物膜的培养多以玻片、细胞爬片、滤膜为载体,可在液体中或固体培养基表面培养,依靠扫描电镜或共聚焦显微镜观察,通常认为观察到多层复合结构即为生物膜结构[5-6]。 生物膜的形成是一个动态过程,先后包括5个步骤[7]:可逆性粘附、不可逆性粘附、早期形成阶段、成熟及消散阶段。虽然很多研究认为支原体培养24小时生物膜即已形成,并以此期生物膜为对象研究其对抗菌素等的抵抗力。但Laura McAuliff等[2]在研究了牛支原体生物膜时有不同的发现。作者利用共聚交显微镜结合SYTO9/PI 荧光探针对牛支原体生物膜形成的动态过程进行观测,发现形成的24及48小时大部分细胞是活的,而通过共聚交显微镜的观察及三维重构发现牛支原体生物膜在最初的24小时仅有一层细胞粘附,48小时才发展成一个非匀质的框架结构,有近20um高,还有通道样结构,此时的生物膜才趋于成熟,同时研究发现培养24小时的牛支原体生物膜对达氟沙星,恩氟沙星,土霉素与游离状态的细胞同样敏感,证明牛支原体培养24小时尚未形成成熟生物膜。可见不同微生物生物膜成熟的时间是存在差异的,在对生物膜特性进行研究之前因先确定其成熟时间点。 支原体生物膜形态与其他微生物相似,可呈网络样、蜂窝状、柱状、蘑菇样、塔样,其间可见水通道,同一种微生物可形成不同结构的生物膜。如肺炎支原体的生物膜最初可形成蜂窝状的区域,在此基础上向外生长成蘑菇状或塔状,塔的直接从小的10um到大于50um,并在塔结构内可见到通道。随着生物膜生长时间的延长,蜂窝状结构中的空洞减少而塔的直径增加,生物膜的形成逐渐趋于成熟[8]。生物膜在不断成熟、丰厚的过程中对内层细胞保护作用不断增强,但由于其深部的细胞营养物质及氧份缺乏也会抑制其生长,正如Laura McAuliff的研究发现培养72小时的生物膜中近70%的细胞都死亡了,活的细胞主要位于生物膜中心。 2影响支原体生物膜形成的因素 生物膜的形成过程中粘附是第一步也是最关键的一步,某些胞外多糖及蛋白质物质是介导粘附的重要基质。如大肠杆菌的表多糖[9],铜绿假单胞菌的藻酸盐[10]等都可促进生物膜的形成。有关支原体的研究也发现支原体的生物膜形成也与某些多糖及蛋白质物质有关。 2.1多糖与生物膜形成 野生型的肺炎支原体可形成一种胞外多糖,即表多糖(exopolysaccharide,EPS)-Ⅰ,它是由当量克分子的葡萄糖
国内外生物信息学发展状况
国内外生物信息学发展状况 1.国外生物信息发展状况 国外非常重视生物信息学的发展各种专业研究机构和公司如雨后春笋般涌现出来,生物科技公司和制药工业内部的生物 信息学部门的数量也与日俱增。美国早在1988年在国会的支持 下就成立了国家生物技术信息中心(NCBI),其目的是进行计 算分子生物学的基础研究,构建和散布分子生物学数据库;欧 洲于1993年3月就着手建立欧洲生物信息学研究所(EBI), 日本也于1995年4月组建了信息生物学中心(CIB)。目前, 绝大部分的核酸和蛋白质数据库由美国、欧洲和日本的3家数 据库系统产生,他们共同组成了 DDBJ/EMBL/Gen Bank国际核 酸序列数据库,每天交换数据,同步更新。以西欧各国为主的 欧洲分子生物学网络组织(EuropeanMolecular Biology Network, EMB Net)是目前国际最大的分子生物信息研究、开 发和服务机构,通过计算机网络使英、德法、瑞士等国生物信 息资源实现共享。在共享网络资源的同时,他们又分别建有自 己的生物信息学机构、二级或更高级的具有各自特色的专业数 据库以及自己的分析技术,服务于本国生物(医学)研究和开 发,有些服务也开放于全世界。 从专业出版业来看,1970年,出现了《Computer Methods and Programs in Biomedicine》这本期刊;到1985年4月, 就有了第一种生物信息学专业期刊《Computer Application
in the Biosciences》。现在,我们可以看到的专业期刊已经很多了。 2 国内生物信息学发展状况 我国生物信息学研究近年来发展较快,相继成立了北京大学生物信息学中心、华大基因组信息学研究中心、中国科学院上海生命科学院生物信息中心,部分高校已经或准备开设生物信息学专业。2002年国家自然科学基金委在生物化学、生物物理学与生物医学工程学学科设立了生物信息学项目,并列入生命科学部优先资助的研究项目。国家 863计划特别设立了生物信息技术主题,从国家需求的层面上推动我国生物信息技术的大力发展[3]。 但是由于起步较晚及诸多原因,我国的生物信息学发展水平远远落后于国外。在PubMed收录的以关键词“Bioinformatics”检索到的历年发表的文章数,可以看出大量的研究文献出现在21世纪以后。其中我国共有138篇占全部5548篇的2.5%,而美国则发表2160篇占全部的39%之多(统计数据截至2004年2月15日)。我国学者在生物信息学领域发表的有高影响力的论文只有不到美国学者发表数量的6%,差距相当大[4]。在生物信息学领域,一些著名院士和教授在各自领域取得了一定成绩,显露出蓬勃发展的势头,有的在国际上还占有一席之地。如北京大学的罗静初和顾孝诚教授在生物信息学网站建设方面、中科院生物物理所的陈润生研究员在EST
生物膜法在污水处理中的研究进展
泉州师范学院 学年论文 论文题目:生物膜法在污水处理中的研究进展指导老师:黄初龙 学院:资源与环境科学学院 专业班级:09级环境工程与管理 学号:090905001 姓名:刘姣
生物膜法在污水处理中的研究进展 摘要:生物膜法在污水处理工艺中是与活性污泥法并行的一种好氧型生物污水处理方法,广泛的应用于工业废水和城市污水处理的二级处理中,也是污水处理的关键环节。与活性污泥法相比,生物膜法具有一些特有优势,比如无需污泥回流,运行管理容易,无污泥膨胀问题,易于微生物生存,运行稳定等。文中简单介绍了生物膜法对磷、氮及一些重金属去除的研究进展。 关键词:生物膜法;污水处理;活性污泥法 Abstract:Biofilm and activated sludge is a parallel-ty pe aerobic biological treatment methods,in the sewage treatment process.They widely used in the secondary treatment of industrial wastewater and urban sewage treatment,and these methods are the key link in sewage treatment.Compared with the activated sludge process,biofilm has some unique advantages.For example,no sludge return,easy operation and management,no sludge expansion,ease of microbial survival,run stable,etc.The paper describes simply biofilm research on the removal of phosphorus,nitrogen and some heavy metals. Key words:B iofilm treatment;sewage treatment;activated sludge 引言 近年来,伴随着经济的快速发展,我国在追求GDP增长的同时也带来一系列的环境问题,其中淡水资源紧缺迫使城镇生活污水处理技术显得尤其重要。然而随着人们生活水平的提高,城镇生活污水中的氮、磷含量增加,有机成分复杂,传统的生物污水处理技术已无法紧随步伐,处理效果不佳,为此,在新型填料的不断开发和完善基础上,生物膜法处理工艺借其处理效率高、剩余污泥产泥量少、运行管理方便等特点得到快速发,在污水处理中有广阔的应用前景。生物膜可认为是由一种或是多种微生物群体组成的,并附着在一种载体表面上进行生长发育[1—2]。 1 生物膜法概述 1.1生物膜法的净水机理 生物膜法和活性污泥法一样都是利用微生物来去除废水中各种有机物的处
浅谈生物信息学在生物方面的应用
浅谈生物信息学在生物方面的应用 生物信息学(bioinformaLics)是以核酸和蛋白质等生物大分子数据库及其相关的图书、文献、资料为主要对象,以数学、信息学、计算机科学为主要手段,对浩如烟海的原始数据和原始资料进行存储、管理、注释、加工,使之成为具有明确生物意义的生物信息。并通过对生物信息的查询、搜索、比较、分析,从中获得基因的编码、凋控、遗传、突变等知识;研究核酸和蛋白质等生物大分子的结构、功能及其相互关系;研究它们在生物体内的物质代谢、能量转移、信息传导等生命活动中的作用机制。 从生物信息学研究的具体内容上看,生物信息学可以用于序列分类、相似性搜索、DNA 序列编码区识别、分子结构与功能预测、进化过程的构建等方面的计算工具已成为变态反应研究工作的重要组成部分。针对核酸序列的分析就是在核酸序列中寻找过敏原基因,找出基因的位置和功能位点的位置,以及标记已知的序列模式等过程。针对蛋白质序列的分析,可以预测出蛋白质的许多物理特性,包括等电点分子量、酶切特性、疏水性、电荷分布等以及蛋白质二级结构预测,三维结构预测等。 生物信息学中的主要方法有:序列比对,结构比对,蛋白质结构的预测,构造分子进化树,聚类等。基因芯片是基因表达谱数据的重要来源。目前生物信息学在基因芯片中的应用主要体现在三个方面。 1、确定芯片检测目标。利用生物信息学方法,查询生物分子信息数据库,取得相应的序列数据,通过序列比对,找出特征序列,作为芯片设计的参照序列。 2、芯片设计。主要包括两个方面,即探针的设计和探针在芯片上的布局,必须根据具体的芯片功能、芯片制备技术采用不同的设计方法。 3、实验数据管理与分析。对基因芯片杂交图像处理,给出实验结果,并运用生物信息学方法对实验进行可靠性分析,得到基因序列变异结果或基因表达分析结果。尽可能将实验结果及分析结果存放在数据库中,将基因芯片数据与公共数据库进行链接,利用数据挖掘方法,揭示各种数据之间的关系。 生物信息学在人类基因组计划中也具有重要的作用。 大规模测序是基因组研究的最基本任务,它的每一个环节都与信息分析紧密相关。目前,从测序仪的光密度采样与分析、碱基读出、载体标识与去除、拼接与组装、填补序列间隙,到重复序列标识、读框预测和基因标注的每一步都是紧密依赖基因组信息学的软件和数据库的。特别是拼接和填补序列间隙更需要把实验设计和信息分析时刻联系在一起.拼接与组装中的难点是处理重复序列,这在含有约30%重复序列的人类基因组中显得尤其突出。 人类基因组的工作草图即将完成,因此发现新基因就成了当务之急。使用基因组信息学的方法通过超大规模计算是发现新基因的重要手段,可以说大部分新基因是靠理论方法预测出来的。比如啤酒酵母完整基因组(约1300万bp)所包含6千多个基因,大约60%是通过信息分析得到的。 当人类基因找到之后,自然要解决的问题是:不同人种间基因有什么差别;正常人和病人基因又有什么差别。”这就是通常所说的SNPs(单核苷酸多态性)。构建SNPs及其相关数据库是基因组研究走向应用的重要步骤。1998年国际已开展了以EST为主发现新Spps 的研究。在我国开展中华民族SNPs研究也是至重要的。总之,生物信息学不仅将赋予人们各种基础研究的重要成果,也会带来巨大的经济效益和社会效益。在未来的几年中DNA 序列数据将以意想不到的速度增长,这更离不开利用生物信息学进行各类数据的分析和解释,研制有效利用和管理数据新工具。生物信息学在功能基因组学同样具有重要的应用目前应用最多的是同源序列比较、模式识别以及蛋白结构预测。所谓同源序列,是指从某一共同祖先经趋异进化而形成的不同序列。利用数据库搜索找出未知核酸或蛋白的同源序列,是序列分析的基础[lol。如利用BLASTn和BLASTx两种软件分别进行核苷酸和氨基
电子版-生物膜动力学的研究现状与展望
生物膜动力学的研究现状与展望 1 引言 生物膜法作为一种高效的废水处理方法,已经在工业界获得了广泛应用。生物膜废水处理系统的性能在很大程度上取决于生物膜的形成及其动力学过程。最近三十年来,各国学者围绕生物膜的形成、发展、结构以及动力学特性等从数学模型、数值模拟和实验研究等方面进行了大量的研究,取得了许多重要进展,为生物膜反应器的设计提供了理论和实验支持,有力地推动了生物膜废水处理工艺的发展。 2 生物膜动力学模型的研究进展 动力学数学模型一直被作为模拟生物膜中微生物动力学行为和生物膜微观结构的一种有力工具,也是将生物膜内微观现象和大规模工艺运行的宏观指标联系起来的关键工具【1】。迄今为止,生物膜动力学数学模型的使用仍在研究领域占主导地位。科研工作者对生物膜形成、构成、结构及功能的兴趣,极大地推动了生物膜动力学数学模型的发展。自20世纪70年代反应-扩散动力学模型提出以来,描述生物膜动力学的模型先后又有Capdeville 增长动力学体系、元胞自动机模型和复合生物膜模型,分别介绍如下: 2.1 反应-扩散动力学模型【2,3】 反应-扩散动力学模型是描述生物膜动力学的最基本的模型。几乎所有的生物膜数学模型都假定生物膜内电子供体、电子受体和所有的营养物质只通过扩散作用传递给微生物(内部传质),而忽略了这些物质从液相主体到生物膜的传递过程(外部传质)。反应-扩散模型将生物膜假设为规则连续介质的稳态膜(包含单一物种),仅考虑一维(1D)物质传输和生化转化作用。生物膜被理想化成具有恒定厚度(f L )和统一细胞密度(f X )的薄膜。从液相主体到生物膜的基质通量是由生物膜内部的微生物活性产生。微生物增长用Monod 方程表示;基质消耗速率(ut r )假定正比于微生物生长速率;基质通量仅用扩散表示。生物膜外部传质限制被认为出现在位于生物膜和液相主体交界面处具有恒定厚度(f L )的边界层中。传质通量采用菲克定律(Fick Law)描述,但其中的扩散系数用有效扩散系数替代:S S e dS J D dx =。这种理想化生物膜的数学模型可以用如下微分方程来表示22?.s S S e f S S S d S q S D X t dx K S ?=-?+,0f x L ≤≤(1) 边界条件为0x =时0S dS dx =(2)f x L =时()S S S e L Sb S dS J D k S S dx ==-(3) 基质利用和扩散由方程(1)描述,边界条件采用式(2)和(3)描述。由于附着表面不可穿透,故此处的通量和基质梯度为零(见式(2))。在生物膜和液相主体交界面处的基质浓度(s S )由质量守恒式确定。即,通过边界层的基质通量必定等于进入生物膜的基质通量(见式(3))。这个理想化的数学模型可以利用有限差分法近似求解。当生物膜处于稳态时,系统可以使用有效因子法和伪解析法求解。关于有效因子法和伪解析法的详细介绍可以参考文献【2,3】。 生物膜反应-扩散理论自20世纪70年代提出后,经过各国学者的大量研究工作而得到完善,并得到了广泛接受和承认。然而,最近十几年来,许多新的实验研究和发现表明,反应.扩散模型的许多假设是过于理想化的,模型的更为合理化是将来研究的重点【4】。 2.2 Capdeville 生物膜增长动力学模型【4,5】 20世纪90年代初,法国CapdeviUe 教授所领导的实验室提出生物膜反应器活性物质和非
生物信息学完整版
一、名词解释 1. 生物信息学: 1)生物信息学包含了生物信息的获取、处理、分析、和解释等在内的一门交叉学科; 2)它综合运用了数学、计算机学和生物学的各种工具来进行研究; 3)目的在于阐明大量生物学数据所包含的生物学意义。 2. BLAST(Basic Local Alignment Search Tool) 直译:基本局部排比搜索工具 意译:基于局部序列排比的常用数据库搜索工具 含义:蛋白质和核酸序列数据库搜索软件系统及相关数据库 3. PSI-BLAST:是一种迭代的搜索方法,可以提高BLAST和FASTA的相似序列发现率。 4. 一致序列:这些序列是指把多序列联配的信息压缩至单条序列,主要的缺点是除了在特 定位置最常见的残基之外,它们不能表示任何概率信息。 5. HMM 隐马尔可夫模型:一种统计模型,它考虑有关匹配、错配和间隔的所有可能的组合 来生成一组序列排列。(课件定义)是蛋白质结构域家族序列的一种严格的统计模型,包括序列的匹配,插入和缺失状态,并根据每种状态的概率分布和状态间的相互转换来生成蛋白质序列。 6. 信息位点:由位点产生的突变数目把其中的一课树与其他树区分开的位点。 7. 非信息位点:对于最大简约法来说没有意义的点。 8. 标度树:分支长度与相邻节点对的差异程度成正比的树。 9. 非标度树:只表示亲缘关系无差异程度信息。 10. 有根树:单一的节点能指派为共同的祖先,从祖先节点只有唯一的路径历经进化到达其 他任何节点。 11. 无根树:只表明节点间的关系,无进化发生方向的信息,通过引入外群或外部参考物种, 可以在无根树中指派根节点。 12. 注释:指从原始序列数据中获得有用的生物学信息。这主要是指在基因组DNA中寻找基 因和其他功能元件(结构注释),并给出这些序列的功能(功能注释)。 13. 聚类分析:一种通过将相似的数据划分到特定的组中以简化大规模数据集的方法。 14. 无监督分析法:这种方法没有内建的分类标准,组的数目和类型只决定于所使用的算法 和数据本身的分析方法。 15. 有监督分析法:这种方法引入某些形式的分类系统,从而将表达模式分配到一个或多个 预定义的类目中。 16. 微阵列芯片:将探针有规律地排列固定于载体上,与标记荧光分子的样品进行杂交,通 过扫描仪扫描对荧光信号的强度进行检测,从而迅速得出所要的信息。 17. 虚拟消化:是基于已知蛋白序列和切断酶的特异性的情况下进行的理论酶切(课件定 义)。是在已知蛋白质序列和蛋白外切酶之类切断试剂的已知特异性的基础上,由计算机进行的一种理论上的蛋白裂解反应。 18. 质谱(MS)是一种准确测定真空中离子的分子质量/电荷比(m/z)的方法,从而使分子质量 的准确确定成为可能。 19. 分子途径是指一组连续起作用以达到共同目标的蛋白质。 20. 虚拟细胞:一种建模手段,把细胞定义为许多结构,分子,反应和物质流的集合体。 21. 先导化合物:是指具有一定药理活性的、可通过结构改造来优化其药理特性而可能导致 药物发现的特殊化合物。就是利用计算机在含有大量化合物三维结构的数据库中,搜索能与生物大分子靶点匹配的化合物,或者搜索能与结合药效团相符的化合物,又称原型物,简称先导物,是通过各种途径或方法得到的具有生物活性的化学结构
结核分枝杆菌生物膜研究进展
Hans Journal of Medicinal Chemistry 药物化学, 2018, 6(3), 78-84 Published Online August 2018 in Hans. https://www.sodocs.net/doc/3619150965.html,/journal/hjmce https://https://www.sodocs.net/doc/3619150965.html,/10.12677/hjmce.2018.63011 Advances on Research of Mycobacterium tuberculosis Biofilms Menglan Gan, Renfeng Wang, Zaichang Yang* School of Pharmacy, Guizhou University, Guiyang Guizhou Received: Aug. 1st, 2018; accepted: Aug. 15th, 2018; published: Aug. 22nd, 2018 Abstract Biofilms refer to a microbial community that is surrounded by a self-generated extracellular po-lymer and attached to the cell surface, but the physiology and genetics definition of the M. tuber-culosis biofilm have not yet been described. Because of its unique physiological state, M. tuberculo-sis biofilms limit the therapeutic effect of anti-tuberculosis drugs, prolong the cycle of tuberculosis treatment, and seriously endanger human health. This article reviewed the formation mechanism, structural composition and related functions and quantitative methods of M. tuberculosis biofilms, and discussed the research ideas of using M. tuberculosis biofilms as novel anti-tuberculosis drugs to shorten the treatment of tuberculosis and provide a new direction for improving the therapeu-tic effect of tuberculosis. Keywords Mycobacterium tuberculosis, Biofilms, Tolerance 结核分枝杆菌生物膜研究进展 甘梦兰,王仁凤,杨再昌* 贵州大学药学院,贵州贵阳 收稿日期:2018年8月1日;录用日期:2018年8月15日;发布日期:2018年8月22日 摘要 生物膜是指被自我产生的细胞外聚合物包裹,并附着在细胞表面的微生物群落,但结核分枝杆菌生物膜*通讯作者。
浅谈生物信息学的发展和前景1
浅谈生物信息学的发展和前景 摘要:生物信息学已成为整个生命科学发展的重要组成部分,成为生命科学研究的前沿。本文对生物信息学的产生背景及其研究现状等方面进行了综述,并展望生物信息学的发展前景。生物信息学的发展在国内、外基本上都处在起步阶段。因此,这是我国生物学赶超世界先进水平的一个百年一遇的极好机会。 关键字:生物信息学、产生背景、发展现状、前景 随着生物科学技术的迅猛发展,生物信息数据资源的增长呈现爆炸之势,同时计算机运算能力的提高和国际互联网络的发展使得对大规模数据的贮存、处理和传输成为可能,为了快捷方便地对已知生物学信息进行科学的组织、有效的管理和进一步分析利用,一门由生命科学和信息科学等多学科相结合特别是由分子生物学与计算机信息处理技术紧密结合而形成的交叉学科——生物信息学(Bioinformatics)应运而生,并大大推动了相关研究的开展, 被誉为“解读生命天书的慧眼”。 一、生物信息学产生的背景 生物信息学是80年代未随着人类基因组计划(Human genome project)的启动而兴起的一门新的交叉学科。它通过对生物学实验数据的获取、加工、存储、检索与分析,进而达到揭示数据所蕴含的生物学意义的目的。由于当前生物信息学发展的主要推动力来自分子生物学,生物信息学的研究主要集中于核苷酸和氨基酸序列的存储、分类、检索和分析等方面,所以目前生物信息学可以狭义地定义为:将计算机科学和数学应用于生物大分子信息的获取、加工、存储、分类、检索与分析,以达到理解这些生物大分子信息的生物学意义的交叉学科。事实上,它是一门理论概念与实践应用并重的学科。 生物信息学的产生发展仅有10年左右的时间---bioinformatics这一名词在1991年左右才在文献中出现,还只是出现在电子出版物的文本中。事实上,生物信息学的存在已有30多年,只不过最初常被称为基因组信息学。美国人类基因组计划中给基因组信息学的定义:它是一个学科领域,包含着基因组信息的获取、处理、存储、分配、分析和解释的所有方面。自1990年美国启动人类基因组计划以来,人与模式生物基因组的测序工作进展极为迅速。迄今已完成了约40多种生物的全基因组测序工作,人基因组约3x109碱基对的测序工作也接近完成。至2000年6月26日,被誉为生命“阿波罗计划”的人类基因组计划终于完成了工作草图,预示着完成人类基因组计划已经指日可待。截止目前为止,仅登录在美国GenBank 数据库中的DNA序列总量已超过70亿碱基对。此外,迄今为止,已有一万多种蛋白质的空间结构以不同的分辨率被测定。基于cDNA序列测序所建立起来的EST数据库其纪录已达数百万条。在这些数据基础上派生、整理出来的数据库已达500余个。这一切构成了一个生物学数据的海洋。这种科学数据的急速和海量积累,在人类的科学研究历史中是空前的。数据并不等于信息和知识,但却是信息和知识的源泉,关键在于如何从中挖掘它们。与正在以指数方式增长的生物学数据相比,人类相关知识的增长(粗略地用每年发表的生物、医学论文数来代表)却十分缓慢。一方面是巨量的数据;另一方面是我们在医学、药物、农业和环保等方面对新知识的渴求,这些新知识将帮助人们改善其生存环境和提高生活质量。这就构成了一个极大的矛盾。这个矛盾就催生了一门新兴的交叉科学,这就是生物信息学。二、生物信息学研究的发展现状 资金和实力非常重要,生物信息的研究投入短期不算大,但是结合成果,其投入相当的大。因为目前生物信息主要在于教学和和研究,商业领域的应用不算很广。如一套LIMS加上软件就要花上数千万。加上相关项目的研究开发,不是国内相关的机构所能承受的。所以需要得到政府的支持和帮助。以及有识之士的投入。否则我们又将远远落后国外。国内的制药行业将永不得翻身!基因的流失(国外一些国家打着给国内免费治疗,分析疾病的考旗帜,
生物信息学发展概况及研究进展
生物信息学发展概况及研究进展 韩龙生物化学与分子生物学2010200531 1 概述 生物信息学是在生命科学、计算机科学和数学的基础上逐步发展而形成的一门新兴的边缘学科,它以核酸和蛋白质为主要研究对象,以数学、计算机科学为主要研究手段,对生物学实验数据进行获取、加工、存储、检索与分析,从而达到揭示数据所蕴含的生物学意义的目的[1]。 生物信息学的发展大致经历了前基因组时代、基因组时代和后基因组时代。目前,它的主要研究内容已经从对DNA和蛋白质序列比较、编码区分析、分子进化转移到大规模的数据整合、可视化,转移到比较基因组学、代谢网络分析、基因表达谱网络分析、蛋白质技术数据分析处理、蛋白质结构与功能分析以及药物靶点筛选等[1]。在后基因组时代的今天,生物信息学已经成为目前极其热门的系统生物学研究的重要手段。 利用各种功能的软件系统平台,目前生物信息学方法主要通过序列比对与分析、功能基因组与基因表达数据的分析、蛋白质结构预测以及基于结构的药物设计等方面应用于各个生命科学研究领域。 1.1序列比对与分析 序列比对是生物信息学的基础,是比较两个或两个以上符号序列的相似性或不相似性。两个序列的比对现在已有较成熟的动态规划算法,以及在此基础上编写的比对软件包——BLAST和FASTA;两个以上序列的多重序列是生物信息学中尚未解决的一个NP完全的组合优化问题,是目前研究的热点[2]。比较经典的算法有SAGA算法[3]、CLUSTAL算法以及隐马尔可夫模型(Hidden Markov Models,HMM)多重序列比对算法,另外,如Notredame等[4]开发的T-Coffee算法、Timo等[5]设计的Kalign算法、张琎等[6]设计的基于GC-GM多序列比对穷举遗传算法,是通过穷举某个特定范围内的所有序列的长度取值,来确定最终最佳比对长度的一种多序列比对算法。这些算法已应用于各种多序列比对软件,并在应用中不断得到优化。 1.2 功能基因组学 在后基因时代的今天,基因组学的研究已从结构基因组学(Structural genomics)转向功能基因组学(Functional genomics)[1] 。功能基因组的任务是进行基因组功能注释(Genome annotation),了解基因功能、认识基因与疾病的关系、掌握基因的产物及其在生命活动中的作用。基因的时空差异表达是功能基因组学研究的理论基础。
生物膜项目可行性研究报告
生物膜项目可行性研究报告 报告摘要 报告通过对项目的市场需求、资源供应、建设规模、工艺路线、设备选型、环境影响、资金筹措、盈利能力等方面的研究调查,在行业专家研究经验的基础上对项目经济效益及社会效益进行科学预测,从而为客户提供全面的、客观的、可靠的项目投资价值评估及项目建设进程等咨询意见。 根据谨慎财务测算,该项目总投资12402.41万元,其中:固定资产投资10496.40万元,占项目总投资的84.63%;流动资金1906.01万元,占项目总投资的15.37%。在固定资产投资中建筑工程投资3342.91万元,占项目总投资的26.95%;设备购置费3292.53万元,占项目总投资的26.55%;其它投资费用3860.96万元,占项目总投资的31.13%。 项目建成投入正常运营后主要生产生物膜产品,根据谨慎财务测算,预期达纲年营业收入15818.00万元,总成本费用11874.62万元,税金及附加68.76万元,利润总额3943.38万元,利税总额4585.11万元,税后净利润2957.53万元,达纲年纳税总额1627.58万元;达纲年投资利润率31.80%,投资利税率36.97%,投资回报率23.85%,全部投资回收期5.69年,提供就业职位286个,达纲年综合节能量24.29吨标准煤/年,项目总节能率26.81%,具有显著的经济效益、社会效益和节能效益。
目录 第一章项目绪论 (7) 一、项目名称及建设性质 (7) 二、项目承办单位 (7) 三、项目建设选址及用地综述 (7) 四、项目土建工程建设指标 (8) 五、设备选型方案 (9) 六、主要能源供应及节能分析 (9) 七、环境保护及清洁生产和安全生产 (9) 八、项目总投资及资金构成 (11) 九、资金筹措方案 (11) 十、项目预期经济效益规划目标 (11) 十一、项目建设进度规划 (12) 十二、综合评价及 (12) 第二章报告编制总体说明 (16) 一、报告编制目的及编制依据 (16) 二、报告编制范围及编制过程 (18) 第三章项目建设背景及必要性 (20) 一、生物膜产业发展规划背景 (20) 二、项目建设背景 (22) 三、项目建设的必要性 (25) 第四章建设规模和产品规划方案合理性分析 (27) 一、建设规模及主要建设内容 (27) 二、产品规划方案及生产纲领 (28)